Виды нуклеотидов в молекуле днк. Нуклеотидный состав и структура ДНК и РНК. Строение структурного звена - нуклеотида
4.2.1. Первичной структурой нуклеиновых кислот называется последовательность расположения мононуклеотидов в цепи ДНК или РНК . Первичная структура нуклеиновых кислот стабилизируется 3",5"-фосфодиэфирными связями. Эти связи образуются при взаимодействии гидроксильной группы в 3"-положении пентозного остатка каждого нуклеотида с фосфатной группой соседнего нуклеотида (рисунок 3.2),
Таким образом, на одном конце полинуклеотидной цепи имеется свободная 5"-фосфатная группа (5"-конец), а на другом - свободная гидроксильная группа в 3"-положении (3"-конец). Нуклеотидные последовательности принято записывать в направлении от 5"-конца к 3"-концу.
Рисунок 4.2. Структура динуклеотида, в состав которого входят аденозин-5"-монофосфат и цитидин-5"-монофосфат.
4.2.2. ДНК (дезоксирибонуклеиновая кислота) содержится в клеточном ядре и имеет молекулярную массу порядка 1011 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, тимин , углевод дезоксирибоза и остатки фосфорной кислоты. Содержание азотистых оснований в молекуле ДНК определяют правила Чаргаффа:
1) количество пуриновых оснований равно количеству пиримидиновых (А + Г = Ц + Т) ;
2) количество аденина и цитозина равно количеству тимина и гуанина соответственно (А = Т; Ц = Г) ;
3) ДНК, выделенные из клеток различных биологических видов, отличаются друг от друга величиной коэффициента специфичности:
(Г + Ц) /(А + Т)
Эти закономерности в строении ДНК объясняются следующими особенностями её вторичной структуры:
1) молекула ДНК построена из двух полинуклеотидных цепей, связанных между собой водородными связями и ориентированных антипараллельно (то есть 3"-конец одной цепи расположен напротив 5"-конца другой цепи и наоборот);
2) водородные связи образуются между комплементарными парами азотистых оснований. Аденину комплементарен тимин; эта пара стабилизируется двумя водородными связями. Гуанину комплементарен цитозин; эта пара стабилизируется тремя водородными связями (см. рисунок б) . Чем больше в молекуле ДНК пар Г-Ц, тем больше её устойчивость к действию высоких температур и ионизирующего излучения;

Рисунок 3.3. Водородные связи между комплементарными азотистыми основаниями.
3) обе цепи ДНК закручены в спираль, имеющую общую ось. Азотистые основания обращены внутрь спирали; кроме водородных, между ними возникают также гидрофобные взаимодействия. Рибозофосфатные части расположены по периферии, образуя остов спирали (см. рисунок 3.4).

Рисунок 3.4. Схема строения ДНК.
4.2.3. РНК (рибонуклеиновая кислота) содержится преимущественно в цитоплазме клетки и имеет молекулярную массу в пределах 104 - 106 Да. В состав её нуклеотидов входят азотистые основания аденин, гуанин, цитозин, урацил , углевод рибоза и остатки фосфорной кислоты. В отличие от ДНК, молекулы РНК построены из одной полинуклеотидной цепи, в которой могут находиться комплементарные друг другу участки (рисунок 3.5). Эти участки могут взаимодействовать между собой, образуя двойные спирали, чередующиеся с неспирализованными участками.

Рисунок 3.5. Схема строения транспортной РНК.
По особенностям структуры и функции различают три основных типа РНК:
1) матричные (информационные) РНК (мРНК) передают информацию о структуре белка из клеточного ядра на рибосомы;
2) транспортные РНК (тРНК) осуществляют транспорт аминокислот к месту синтеза белка;
3) рибосомальные РНК (рРНК) входят в состав рибосом, участвуют в синтезе белка.
Все живое на планете состоит из множества клеток, поддерживающих упорядоченность своей организации за счет содержащейся в ядре генетической информации. Она сохраняется, реализуется и передается сложными высокомолекулярными соединениями - нуклеиновыми кислотами, состоящими из мономерных звеньев - нуклеотидов. Роль нуклеиновых кислот невозможно переоценить. Стабильностью их структуры определяется нормальная жизнедеятельность организма, а любые отклонения в строении неминуемо приводят к изменению клеточной организации, активности физиологических процессов и жизнеспособности клеток в целом.
Понятие нуклеотида и его свойства
Каждая или РНК собрана из более мелких мономерных соединений - нуклеотидов. Другими словами, нуклеотид - это строительный материал для нуклеиновых кислот, коферментов и многих других биологических соединений, которые крайне необходимы клетке в процессе ее жизнедеятельности.
К основным свойствам этих незаменимых веществ можно отнести:
Хранение информации о и наследуемых признаках;
. осуществление контроля над ростом и репродукцией;
. участие в метаболизме и многих других физиологических процессах, протекающих в клетке.
Говоря о нуклеотидах, нельзя не остановиться на таком важном вопросе, как их структура и состав.

Каждый нуклеотид состоит из:
Сахарного остатка;
. азотистого основания;
. фосфатной группы или остатка фосфорной кислоты.
Можно сказать, что нуклеотид - это сложное органическое соединение. В зависимости от видового состава азотистых оснований и типа пентозы в структуре нуклеотида нуклеиновые кислоты подразделяются на:
Дезоксирибонуклеиновую кислоту, или ДНК;
. рибонуклеиновую кислоту, или РНК.
Состав нуклеиновых кислот
В нуклеиновых кислотах сахар представлен пентозой. Это пятиуглеродный сахар, в ДНК его называют дезоксирибозой, в РНК - рибозой. Каждая молекула пентозы имеет пять атомов углерода, четыре из них вместе с атомом кислорода образуют пятичленное кольцо, а пятый входит в группу НО-СН2.
Положение каждого атома углерода в молекуле пентозы обозначается арабской цифрой со штрихом (1C´, 2C´, 3C´, 4C´, 5C´). Поскольку все процессы считывания с молекулы нуклеиновой кислоты имеют строгую направленность, нумерация атомов углерода и их расположение в кольце служат своего рода указателем правильного направления.
По гидроксильной группе к третьему и пятому углеродным атомам (3С´ и 5С´) присоединен остаток фосфорной кислоты. Он и определяет химическую принадлежность ДНК и РНК к группе кислот.
К первому углеродному атому (1С´) в молекуле сахара присоединено азотистое основание.
Видовой состав азотистых оснований
Нуклеотиды ДНК по азотистому основанию представлены четырьмя видами:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. тимином (Т).
Первые два относятся к классу пуринов, два последних - пиримидинов. По молекулярной массе пуриновые всегда тяжелее пиримидиновых.

Нуклеотиды РНК по азотистому основанию представлены:
Аденином (А);
. гуанином (Г);
. цитозином (Ц);
. урацилом (У).
Урацил так же, как и тимин, является пиримидиновым основанием.
В научной литературе нередко можно встретить и другое обозначение азотистых оснований - латинскими буквами (A, T, C, G, U).
Подробнее остановимся на химической структуре пуринов и пиримидинов.

Пиримидины, а именно цитозин, тимин и урацил, в своем составе представлены двумя атомами азота и четырьмя атомами углерода, образующих шестичленное кольцо. Каждый атом имеет свой номер от 1 до 6.
Пурины (аденин и гуанин) состоят из пиримидина и имидазола или двух гетероциклов. Молекула пуриновых оснований представлена четырьмя атомами азота и пятью атомами углерода. Каждый атом пронумерован от 1 до 9.
В результате соединения азотистого основания и остатка пентозы образуется нуклеозид. Нуклеотид - это соединение нуклеозида и фосфатной группы.
Образование фосфодиэфирных связей
Важно разобраться в вопросе о том, как соединяются нуклеотиды в полипептидную цепь и образуют молекулу нуклеиновой кислоты. Происходит это за счет так называемых фосфодиэфирных связей.
Взаимодействие двух нуклеотидов дает динуклеотид. Образование нового соединения происходит путем конденсации, когда между фосфатным остатком одного мономера и гидроксигруппой пентозы другого возникает фосфодиэфирная связь.
Синтез полинуклеотида - неоднократное повторение этой реакции (несколько миллионов раз). Полинуклеотидная цепь строится посредством образования фосфодиэфирных связей между третьим и пятым углеродами сахаров (3С´ и 5С´).
Сборка полинуклеотида - сложный процесс, протекающий при участии фермента ДНК-полимеразы, которая обеспечивает рост цепи только с одного конца (3´) со свободной гидроксигруппой.
Структура молекулы ДНК
Молекула ДНК, так же как и белка, может иметь первичную, вторичную и третичную структуру.

Последовательность нуклеотидов в цепи ДНК определяет ее первичную формируется за счет водородных связей, в основе возникновения которых положен принцип комплементарности. Другими словами, при синтезе двойной действует определенная закономерность: аденин одной цепи соответствует тимину другой, гуанин - цитозину, и наоборот. Пары аденина и тимина или гуанина и цитозина образуются за счет двух в первом и трех в последнем случае водородных связей. Такое соединение нуклеотидов обеспечивает прочную связь цепей и равное расстояние между ними.
Зная последовательность нуклеотидов одной цепи ДНК, по принципу комплементарности или дополнения можно достроить вторую.
Третичная структура ДНК образована за счет сложных трехмерных связей, что делает ее молекулу более компактной и способной размещаться в малом объеме клетки. Так, например, длина ДНК кишечной палочки составляет более 1 мм, тогда как длина клетки - меньше 5 мкм.
Число нуклеотидов в ДНК, а именно их количественное соотношение, подчиняется правилу Чергаффа (число пуриновых оснований всегда равно количеству пиримидиновых). Расстояние между нуклеотидами - величина постоянная, равная 0,34 нм, как и их молекулярная масса.
Структура молекулы РНК
РНК представлена одной полинуклеотидной цепочкой, образованной через между пентозой (в данном случае рибозой) и фосфатным остатком. По длине она значительно короче ДНК. По видовому составу азотистых оснований в нуклеотиде также имеются различия. В РНК вместо пиримидинового основания тимина используется урацил. В зависимости от функций, выполняемых в организме, РНК может быть трех типов.

Рибосомальная (рРНК) - содержит обычно от 3000 до 5000 нуклеотидов. Как необходимый структурный компонент принимает участие в формировании активного центра рибосом, места осуществления одного из важнейших процессов в клетке — биосинтеза белка.
. Транспортная (тРНК) - состоит в среднем из 75 - 95 нуклеотидов, осуществляет перенос нужной аминокислоты к месту синтеза полипептида в рибосоме. Каждый вид тРНК (не менее 40) имеет свою, присущую только ему последовательность мономеров или нуклеотидов.
. Информационная (иРНК) - по нуклеотидному составу весьма разнообразна. Переносит генетическую информацию от ДНК к рибосомам, выступает в роли матрицы для синтеза белковой молекулы.
Роль нуклеотидов в организме
Нуклеотиды в клетке выполняют ряд важнейших функций:
Используются в качестве структурных блоков для нуклеиновых кислот (нуклеотиды пуринового и пиримидинового рядов);
. участвуют во многих обменных процессах в клетке;
. входят в состав АТФ - главного источника энергии в клетках;
. выступают в роли переносчиков восстановительных эквивалентов в клетках (НАД+, НАДФ+, ФАД, ФМН);
. выполняют функцию биорегуляторов;
. могут рассматриваться как вторые вестники внеклеточного регулярного синтеза (например, цАМФ или цГМФ).
Нуклеотид - это мономерная единица, образующая более сложные соединения - нуклеиновые кислоты, без которых невозможна передача генетической информации, ее хранение и воспроизведение. Свободные нуклеотиды являются главными компонентами, участвующими в сигнальных и энергетических процессах, поддерживающих нормальную жизнедеятельность клеток и организма в целом.
Нуклеиновые кислоты - это природные высокомолекулярные соединения (полинуклеотиды), которые играют огромную роль в хранении и передаче наследственной информации в живых организмах.
Молекулярная масса нуклеиновых кислот может меняться от сотен тысяч до десятков миллиардов. Они были открыты и выделены из клеточных ядер еще в XIX в., однако их биологическая роль была выяснена только во второй половине XX в.
В состав нуклеотида - структурного звена нуклеиновых кислот - входят три составные части:
1) азотистое основание - пиримидиновое или пуриновое
Пиримидиновые основания – производные пиримидина, входящие в состав нуклеиновых кислот: урацил, тимин, цитозин .
Для оснований, содержащих группу –ОН, характерно подвижное равновесие структурных изомеров, обусловленное переносом протона от кислорода к азоту и наоборот:
Пуриновые основания - производные пурина, входящие в состав нуклеиновых кислот: аденин, гуанин .
Гуанин существует в виде двух структурных изомеров:
2) моносахарид
Рибоза и 2-дезоксирибоза относятся к моносахаридам, содержащим пять углеродных атомов. В состав нуклеиновых кислот они входят в циклических β-формах:
3) остаток фосфорной кислоты
ДНК и РНК
В зависимости от того, какой моносахарид содержится в структурном звене полинуклеотида - рибоза или 2-дезоксирибоза , различают
· рибонуклеиновые кислоты (РНК) и
· дезоксирибонуклеиновые кислоты (ДНК)
В
главную (сахарофосфатную) цепь РНК входят остатки рибозы
, а в ДНК – 2-дезоксирибозы
.
Нуклеотидные звенья макромолекул ДНК могут содержать аденин, гуанин, цитозин
и тимин
. Состав РНК отличается тем, что вместо тимина
присутствует
урацил
.
Молекулярная масса ДНК достигает десятков миллионов а.е.м. Это самые длинные из известных макромолекул. Значительно меньше молекулярная масса РНК (от нескольких сотен до десятков тысяч). ДНК содержатся в основном в ядрах клеток, РНК – в рибосомах и протоплазме клеток.
При описании строения нуклеиновых кислот учитывают различные уровни организации макромолекул: первичную и вторичную структуру.
· Первичная структура нуклеиновых кислот – это нуклеотидный состав и определенная последовательность нуклеотидных звеньев в полимерной цепи.
Например:
В сокращённом однобуквенном обозначении эта структура записывается как
...– А – Г – Ц –...
· Под вторичной структурой нуклеиновых кислот понимают пространственно упорядоченные формы полинуклеотидных цепей.
Вторичная структура ДНК представляет собой две параллельные неразветвленные полинуклеотидные цепи, закрученные вокруг общей оси в двойную спираль.
Такая пространственная структура удерживается множеством водородных связей, образуемых азотистыми основаниями, направленными внутрь спирали.Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи. Эти основания составляют комплементарные пары (от лат. complementum - дополнение).
Образование водородных связей между комплементарными парами оснований обусловлено их пространственным соответствием.
Пиримидиновое основание комплементарно пуриновому основанию:
Водородные связи между другими парами оснований не позволяют им разместиться в структуре двойной спирали. Таким образом,
· ТИМИН (Т) комплементарен АДЕНИНУ (А),
· ЦИТОЗИН (Ц) комплементарен ГУАНИНУ (Г).
Комплементарность оснований определяет комплементарность цепей в молекулах ДНК.
Комплементарность полинуклеотидных цепей служит химической основой главной функции ДНК – хранения и передачи наследственных признаков.
Способность ДНК не только хранить, но и использовать генетическую информацию определяется следующими ее свойствами:
· молекулы ДНК способны к репликации (удвоению), т.е. могут обеспечить возможность синтеза других молекул ДНК, идентичных исходным, поскольку последовательность оснований в одной из цепей двойной спирали контролирует их расположение в другой цепи.
· молекулы ДНК могут направлять совершенно точным и определенным образом синтез белков, специфичных для организмов данного вида.
Вторичная структура РНК
В отличие от ДНК, молекулы РНК состоят из одной полинуклеотидной цепи и не имеют строго определенной пространственной формы (вторичная структура РНК зависит от их биологических функций).
Основная роль РНК – непосредственное участие в биосинтезе белка.
Известны три вида клеточных РНК, которые отличаются по местоположению в клетке, составу, размерам и свойствам, определяющим их специфическую роль в образовании белковых макромолекул:
· информационные (матричные) РНК передают закодированную в ДНК информацию о структуре белка от ядра клетки к рибосомам, где и осуществляется синтез белка;
· транспортные РНК собирают аминокислоты в цитоплазме клетки и переносят их в рибосому; молекулы РНК этого типа "узнают" по соответствующим участкам цепи информационной РНК, какие аминокислоты должны участвовать в синтезе белка;
·
рибосомные
РНК обеспечивают синтез белка определенного строения, считывая информацию с
информационной (матричной) РНК.
Нуклеотидный состав, т.е. набор и соотношение нуклеотидных компонентов, служит очень важной характеристикой нуклеиновых кислот. Один из основных путей установления состава нуклеиновых кислот основан на исследовании продуктов их гидролитического расщепления. Поскольку межнуклеотидные связи в полинуклеотидах являются сложноэфирными, то полинуклеотидные цепи способны гидролизоваться как в кислой, так и щелочной среде.
Химический гидролиз ДНК почти не используется из-за осложнения его побочными процессами. Более предпочтителен ферментативный гидролиз ДНК под действием нуклеаз. Обычно для этой цели используют змеиный яд, в котором содержатся ферменты, расщепляющие сложноэфирную связь с фосфорной кислотой (фосфодиэстеразы и фосфомоноэстеразы). Нуклеазы проявляют специфичность по отношению к типу нуклеиновых кислот; их делят на рибонуклеазы и дезоксирибонуклезы.
Выделение и идентификацию компонентов нуклеиновых кислот производят с помощью физико-химических методов. Очень важную роль в разделении сложных смесей играют хроматографические методы. Пиримидиновые и пуриновые основания, обладающие вследствие ароматического характера заметным поглощением около 260 нм, обычно идентифицируют с помощью УФ-спектроскопии. Поскольку нуклеотиды имеют кислотный характер и способны находиться в ионизированном состоянии, то для их идентификации используют также электрофорез.
Наряду с определением нуклеотидного состава важнейшая задача состоит и в установлении нуклеотидной последовательности, т.е. порядка чередования нуклеотидных звеньев. Общий подход заключается в использовании блочного метода: сначала полинуклеотидную цепь направленно расщепляют на более мелкие блоки – олигомеры и определяют в них нуклеотидную последовательность. Такой анализ повторяют дважды, используя во второй раз такие расщепляющие агенты, которые делят цель на фрагменты в иных местах по сравнению с первым разом. Полинуклеотидную цепь расщепляют на довольно короткие фрагменты. Более длинные олигонуклеотиды пока еще трудно поддаются изучению.
Первичная структура нуклеиновых кислот определяется природой и последовательностью нуклеотидных звеньев, связанных сложноэфирными связями между пентозами и фосфатными группами (рис 13).
Рис. 13. Первичная структура участка цепи нуклеиновых кислот
В составе молекулы ДНК выделено значительно большее число нуклеотидных остатков, чем в молекуле РНК. Молекулярная масса ДНК порядка 10 млн; ДНК в условиях клетки нерастворима. Длина молекул ДНК человека составляет примерно 3 - 5 см; молекула РНК значительно короче - менее 0,01 см.
Вторичная структура нуклеиновых кислот. Согласно вторичной структуре полинуклеотидная цепь ДНК представляет собой двойную спираль, в которой пуриновые и пиримидиновые основания направлены внутрь. Между пуриновым основаниями одной цепи и пиримидиновым основанием другой цепи имеются водородные связи, стабилизирующие такую структуру. Основания, образующие пары, связанные водородными связями,называются комплементарными . В ДНК комплементарными будут: аденин – тимин, образующие между собой две водородные связи, и гуанин – цитозин, связанные тремя водородными связями (рис 14). Это означает, что пуриновым основаниям аденину и гуанину в одной цепи будут соответствовать пиримидиновые основания тимин и цитозин в другой цепи. Полинуклеотидные цепи, образующие двойную спираль, не идентичны, но комплементарны между собой.
Рис. 14. Водородные связи в паре оснований гуанин -цитозин (а), аденин – тимин (б)
Макромолекулы ДНК связаны между собой попарно при помощи водородных связей в виде двойной спирали постоянного диаметра (рис. 15). Остатки нуклеиновых оснований направлены внутрь спирали, диаметр которой равен примерно 2 нм.
На один виток спирали приходится 10 пар оснований. Для обеспечения наибольшей устойчивости этой структуры водородных связей должно быть максимально много. Только при выполнении этого условия обеспечивается экспериментально доказанное постоянство суммарных размеров боковых групп и неизменность диаметра двойной спирали на всем ее протяжении. В этой взаимной обусловленности последовательности звеньев в обеих цепях заключается принцип комплементарности.
Комплементарность цепей и последовательность звеньев составляют химическую основу важнейших функций нуклеиновых кислот: ДНК - хранение и передача наследственной информации, а РНК - непосредственное участие в биосинтезе белка. Молекулярная масса ДНК варьирует от нескольких миллионов до десятка миллиардов, у РНК - от десятка тысяч до нескольких миллионов.
Комплементарность оснований лежит в основе закономерностей, сформулированных Э. Чаргаффом, которым подчиняется нуклеотидный состав ДНК различного происхождения.
Правила Чаргаффа:
1)  количество пуриновых оснований равно количеству пиримидиновых оснований, т.е. (А+Г)=(Ц+Т).
количество пуриновых оснований равно количеству пиримидиновых оснований, т.е. (А+Г)=(Ц+Т).
2) Количество аденина равно количеству тимина (А=Т); аналогично количество гуанина равно количеству цитозина (Г=Ц).
3) Количество оснований, содержащих аминогруппу в положении 4 пиримидинового и положении 6 пуринового ядра, равно количеству оснований, содержащих в этих же положениях оксогруппу. Это означает, что А+Ц=Г+Т.
Для РНК правила Чаргаффа либо не выполняются, либо выполняются с некоторым приближением. Это обусловлено тем, что в составе РНК содержится много минорных оснований.
Сравнение макромолекулы ДНК с винтовой лестницей наводит на мысль об ее хиральности. Действительно, природные ДНК обладают оптической активностью. В то же время смеси нуклеотидов, составляющих ДНК, а также разупорядоченные полинуклеотические цепи оптически неактивны. Это свидетельствует о том, что оптическая активность природных ДНК связана с хиральностью их вторичной структуры.
Каркас спирали образован чередующимися углеводными и фосфатными остатками. Окружающая водная среда контактирует с гидрофильной частью спирали, а внутренняя часть спирали (основания) с водой не контактирует.
Молекула ДНК, в отличие от молекулы РНК, в большинстве случаев состоит из двух комплементарных взаимозакрученных цепей. В зависимости от длины витка и угла спирали, а также ряда других ее геометрических параметров, различают, более десяти разнообразных упорядоченных спиральных структур ДНК. В стабилизации этих структур наряду с водородными связями, действующими поперек спирали, большую роль играют межмолекулярные взаимодействия, направленные вдоль спирали между соседними пространственно сближенными азотистыми основаниями. Поскольку эти взаимодействия направлены вдоль стопки азотистых оснований молекулы ДНК, их называют стэкинг-взаимодействиями. Таким образом, взаимодействия азотистых оснований между собой скрепляют двойную спираль молекулы ДНК и вдоль, и поперек ее оси.
Сильное стэкинг-взаимодействие всегда усиливает водородные связи между основаниями, способствуя уплотнению спирали. Вследствие этого молекулы воды из окружающего раствора связываются в основном с пентозофосфатным остовом ДНК, полярные группы которого находятся на поверхности спирали. При ослаблении стэкинг-взаимодействия молекулы воды, проникая внутрь спирали, конкурентно взаимодействуют с полярными группами оснований, инициируют дестабилизацию и способствуют дальнейшему распаду двойной спирали. Все это свидетельствует о динамичности вторичной структуры ДНК под воздействием компонентов окружающего раствора. Двойная спираль характерна для большинства молекул ДНК. Однако ДНК может иметь и другие формы. В некоторых вирусах содержится одноцепочечная ДНК, встречаются также кольцевые формы.
Биспиральные структуры в молекулах РНК возникают в пределах одной и той же цепи в тех зонах, где расположены комплементарные азотистые основания аденин - урацил и гуанин - цитозин (рис. 16). В результате вторичная структура молекулы РНК содержит биспиральные участки и петли, число и размеры которых определяются первичной структурой молекулы и составом окружающего раствора.
Рис. 16. Вторичная структура молекулы РНК
Третичная структура нуклеиновых кислот. Двойная спираль молекул ДНК существует в виде линейной, кольцевой, суперкольцевой и компактных клубковых форм. Между этими формами совершаются взаимные переходы при действии особой группы ферментов – топоизомераз, изменяющих пространственную структуру (рис 17).
Рис. 17. Третичная структура молекулы ДНК:
а -линейная, б - кольцевая, в - суперкольцевая, г - компактный клубок
Третичная структура многих молекул РНК пока еще требует окончательного выяснения, но уже установлено, что она зависит не только от первичной и вторичной структуры, но и от состава окружающего раствора.
- это сложные мономеры, из которых собраны гетерополимерные молекулы. ДНК и РНК. Свободные нуклеотиды участвуют в сигнальных и энергетических процессах жизнедеятельности. ДНК-нуклеотиды и РНК-нуклеотиды имеют общий план строения, но различаются по строению сахара-пентозы. В ДНК-нуклеотидах используется сахар дезоксирибоза, а в РНК-нуклеотидах - рибоза.
Структура нуклеотида
В каждом нуклеотиде можно выделить 3 части:
1. Углевод - это пятичленный сахар-пентоза (рибоза или дезоксирибоза).
2. Фосфорный остаток (фосфат) - это остаток фосфорной кислоты.
3. Азотистое основание - это соединение, в котором много атомов азота. В нуклеиновых кислотах используется всего 5 видов азотистых оснований: Аденин, Тимин, Гуанин, Цитозин, Урацил. В ДНК - 4 вида: Аденин, Тимин, Гуанин, Цитозин. В РНК - тоже 4 вида: Аденин, Урацил, Гуанин, Цитозин, Легко заметить, что в РНК происходит замещение Тимина на Урацил по сравнению с ДНК.
Общая структурная формула пентозы (рибозы или дезоксирибозы), молекулы которой образуют "скелет" нуклеиновых кислот:
Если Х заменить на Н (Х = Н) - то получаются дезоксирибонуклеозиды; если Х заменить на ОН (Х = ОН) - то получаются рибонуклеозиды. Если вместо R подставить азотистое основание (пуриновое или пиримидиновое) - то получится конкретный нуклеотид.
Важно обратить внимание на те положения атомов углерода в пентозе, которые обозначены как 3" и 5". Нумерация атомов углерода начинается от атома кислорода вверху и идёт по часовой стрелке. Последним получается атом углерода (5"), который располагается за пределами пентозного кольца и образует, можно сказать, "хвостик" у пентозы. Так вот, при наращивании цепочки из нуклеотидов фермент может присоединить новый нуклеотид только к углероду 3" и ни к какому другому. Поэтому 5"-конец нуклеотидной цепочки никогда не сможет иметь продолжения, удлинняться может только 3"-конец.



Сравните нуклеотид для РНК с нуклеотидом для ДНК.
Попробуйте узнать, какой это нуклеотид, в таком представлении:

АТФ - свободный нуклеотид
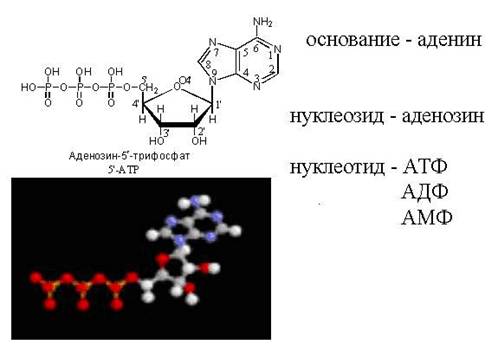
цАМФ - "закольцованная" молекула АТФ

Схема строения нуклеотида

Обратите внимание на то, что активированный нуклеотид, способный наращивать цепочку ДНК или РНК, имеет "трифосфатный хвостик". Именно этим "энергонасыщенным" хвостиком он может присоединиться к уже имеющейся цепочке растущей нуклеиновой кислоты. Фосфатный хвостик сидит на 5-м атоме углерода, так что это положение углерода уже занято фосфатами и предназнено для прикрепления. К чему же его прикрепить? Только к углероду в положении 3". После прикрепления данный нуклеотид сам станет мишенью дла прикрепления следующего нуклеотида. "Принимающая сторона" предоставляет углерод в положении 3", а "прибывающая сторона" цепляется к нему фосфатным хвостиком, находящимся в положении 5". В целом цепочка растёт со стороны 3".
Наращивание нуклеотидной цепочки ДНК
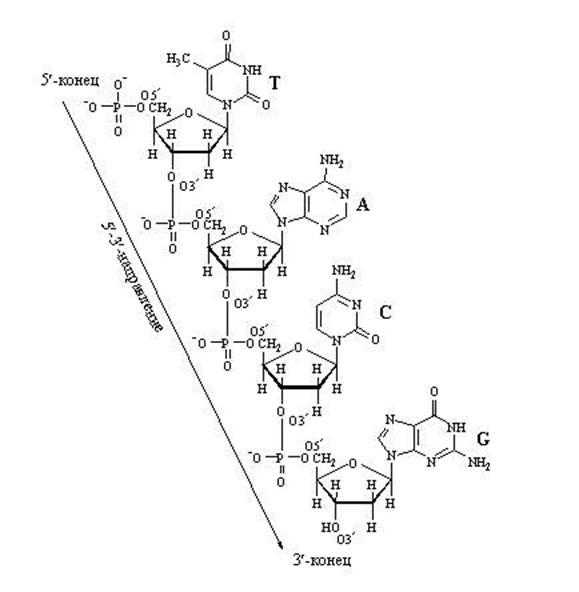
Наращивание цепочки за счёт "продольных" связей между нуклеотидами может идти только в одном направлении: от 5" ⇒ к 3", т.к. новый нуклеотид можно присоединить только к 3"-концу цепочки, но не к 5"-концу.
Пары нуклеотидов, связанные "поперечными" комплементарными связями своих азотистых оснований

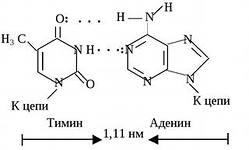
Участок двойной спирали ДНК

Найдите признаки антипараллельности двух цепей ДНК.
Найдите пары нуклеотидов с двойными и тройными комплементарными связями.