Предмет, задачи и цели молекулярной биологии. Биохимия и молекулярная биология - где учиться? Что изучает молекулярная биология кратко
Успехи в изучении нуклеиновых кислот и биосинтеза белка привели к созданию ряда методов, имеющих большое прикладное значение в медицине, сельском хозяйстве и ряде других отраслей.
После того, как был изучен генетический код и основные принципы хранения и реализации наследственной информации, развитие молекулярной биологии зашло в тупик, так как не было методов, которые позволяли манипулировать генами, выделять и изменять их. Появление этих методов произошло в 1970-1980х годах. Это дало мощный толчок развитию этой области науки, которая и сегодня переживает период расцвета. Прежде всего, эти методы касаются получения индивидуальных генов и их введения в клетки других организмов (молекулярное клонирование и трансгенез, ПЦР), а также методов определения последовательности нуклеотидов в генах (секвенирования ДНК и РНК). Ниже эти методы будут рассмотрены более подробно. Мы начнем с простейшего базового метода - электрофореза и затем перейдем к более сложным методам.
ЭЛЕКТРОФОРЕЗ ДНК
Это базовый метод работы с ДНК, применяющийся вместе с практическими всеми другими методами для выделения нужных молекул и анализа результатов. Для разделения фрагментов ДНК по длине применяется метод электрофореза в геле. ДНК - кислота, ее молекулы содержат остатки фосфорной кислоты, которые отщепляют протон и приобретают отрицательный заряд (рис. 1).
Поэтому в электрическом поле молекулы ДНК движутся к аноду - положительно заряженному электроду. Это происходит в растворе электролитов, содержащем ионы-носители заряда, благодаря чему этот раствор проводит ток. Чтобы разделить фрагменты, применяется плотный гель из полимеров (агарозы либо полиакриламида). Молекулы ДНК "запутываются" в нем тем больше, чем они длиннее, и поэтому наиболее длинные молекулы движутся медленнее всего, а наиболее короткие - быстрее всего (рис. 2). Заблаговременно или после электрофореза гель обрабатывают красителями, связывающимися с ДНК и флуоресцирующими в ультрафиолетовом свете, и получают картину полос в геле (см. рис. 3). Для определения длин фрагментов ДНК образца их сравнивают с маркером - набором фрагментов стандартных длин, нанесенных параллельно на тот же гель (рис. 4).



Важнейшими инструментами для работы с ДНК являются ферменты, осуществляющие превращения ДНК в живых клетках: ДНК-полимеразы, ДНК-лигазы и рестрикционные эндонуклеазы, или рестриктазы. ДНК-полимеразы осуществляют матричный синтез ДНК, что позволяет размножать ДНК в пробирке. ДНК-лигазы сшивают между собой молекулы ДНК или залечивают бреши них. Рестрикционные эндонуклеазы , или рестриктазы , разрезают молекулы ДНК по строго определённым последовательностям, что позволяет вырезать отдельные фрагменты из общей массы ДНК. Эти фрагменты могут в каких-то случаях содержать отдельные гены.
рестриктазы
Последовательности, узнаваемые рестриктазами, симметричны, и разрывы могут происходить в середине такой последовательности или со сдвигом (в одном и том же месте в обеих нитях ДНК). Схема действия разных типов рестриктаз показана на рис. 1. В первом случае получаются так называемые «тупые» концы, а во втором – «липкие» концы. В случае «липких» концов дна цепь оказывается короче другой, образуется однонитевой участок с симметричной последовательностью, одинаковой на обоих образующихся концах.
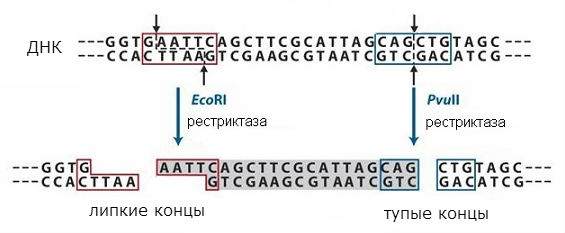
Концевые последовательности будут одинаковыми при расщеплении любой ДНК данной рестриктазой и могут снова соединяться, так как имеют комплементарные последовательности. Их можно сшить с помощью ДНК-лигазы и получить единую молекулу. Таким образом удаётся объединить фрагменты двух разных ДНК и получить так называемые рекомбинантные ДНК . Этот подход используется в методе молекулярного клонирования, позволяющего получить индивидуальные гены и ввести их в клетки, которые могут образовывать закодированный в гене белок.
молекулярное клонирование
В молекулярном клонировании используется две молекулы ДНК - вставка, содержащая интересующий ген, и вектор - ДНК, выступающая в роли носителя. Вставку "вшивают" в вектор пр помощи ферментов, получая новую, рекомбинантную молекулу ДНК, затем эту молекулу внедряют в клетки-хозяева, и эти клетки образуют колонии на питательной среде. Колония - это потомство одной клетки, то есть клон, все клетки колонии генетически идентичны и содержат одну и ту же рекомбинантную ДНК. Отсюда термин "молекулярное клонирование", то есть получение клона клеток, содержащих интересующий нас фрагмент ДНК. После того, как колонии, содержащие интересующую нас вставку, получены, можно различными методами характеризовать эту вставку, например, определить ее точную последовательность. Также клетки могут производить кодируемый вставкой белок, если она содержит функциональный ген.
При внедрении рекомбинантной молекулы в клетки происходит генетическая трансформация этих клеток. Трансформация - процесс поглощения клеткой организма свободной молекулы ДНК из среды и встраивания её в геном, что приводит к появлению у такой клетки новых для неё наследуемых признаков, характерных для организма-донора ДНК. Например, если встраиваемая молекула содержит ген устойчивости к антибиотику ампициллину, то трансформированные бактерии будут расти в его присутствии. До трансформации ампициллин вызывал их гибель, то есть у трансформированных клеток возникает новый признак.
ВЕКТОРЫ
Вектор должен обладать рядом свойств:
Во-первых, это относительно небольшая молекула ДНК, чтобы ей было легко манипулировать.
Во-вторых, для того, чтобы ДНК сохранялась и размножалась в клетке, она должна содержать определённую последовательность, обеспечивающую её репликацию (точку начала репликации, или origin of replication).
В-третьих, она должна содержать ген-маркер , который обеспечивает отбор только тех клеток, в которые попал вектор. Обычно это гены устойчивости к антибиотикам - тогда в присутствии антибиотика все не содержащие вектора клетки погибают.
Клонирование генов чаще всего проводят в клетках бактерий, так как они просты в культивировании и быстро размножаются. В клетке бактерии обычно присутствует одна большая кольцевая молекула ДНК, длиной в несколько миллионов пар нуклеотидов, содержащая все необходимые бактерии гены - бактериальная хромосома. Кроме неё в некоторых бактериях существуют небольшие (несколько тысяч пар нуклеотидов) кольцевые ДНК, называемые плазмидами (рис. 2). Они, также как и основная ДНК, содержат последовательность нуклеотидов, обеспечивающую способность ДНК реплицироваться (ori). Плазмиды реплицируются независимо от основной (хромосомной) ДНК, поэтому присутствуют в клетке в большом количестве копий. Многие из таких плазмид несут гены устойчивости к антибиотикам, что позволяет отличить клетки, несущие плазмиду, от обычных клеток. Чаще используются плазмиды, несущие два гена, обеспечивающие устойчивость к двум антибиотикам, например, к тетрациклину и апмицилину. Существуют простые методы выделения таких плазмидных ДНК, свободных от ДНК основной хромосомы бактерии.

ЗНАЧЕНИЕ ТРАНСГЕНЕЗА
Перенос генов из одного организма в другой называют трансгенезом , а такие модифицированные организмы - трансгенными . Методом переноса генов в клетки микроорганизмов получают рекомбинантные белковые препараты для нужд медицины, в частности, человеческие белки, не вызывающие иммунного отторжения - интерфероны, инсулин и другие белковые гормоны, клеточные факторы роста, а также белки для производства вакцин. В более сложных случаях, когда модификация белков проходит правильно только в клетках эукариот, применяют трансгенные клеточные культуры или трансгенных животных, в частности, скот (прежде всего коз), который выделяет необходимые белки в молоко, или же белки выделяют из их крови. Так получают антитела, факторы свертывания крови и другие белки. Методом трансгенеза получают культурные растения, устойчивые к гербицидам и вредителям и обладающие другими полезными свойствами. При помощью трансгенных микроорганизмов очищают сточные воды и борются с загрязнениями, существуют даже трансгенные микробы, которые могут расщеплять нефть. Помимо этого, трансгенные технологии незаменимы в научных исследованиях - развитие биологии сегодня немыслимо без рутинного применения методов модификации и переноса генов.
технология молекулярного клонирования
вставки
Для получения индивидуального гена из какого-либо организма из него выделяют всю хромосомную ДНК и расщепляют её одной или двумя рестриктазами. Ферменты подбирают так, чтобы они не разрезали интересующий нас ген, а делали разрывы по его краям, а в плазмидной ДНК делали 1 разрыв в одном из генов устойчивости, например, к ампицилину.
Процесс молекулярного клонирования включает следующие этапы:
Разрезание и сшивание - конструирование из вставки и вектора единой рекомбинантной молекулы.
Трансформация - внедрение рекомбинантной молекулы в клетки.
Селекция - отбор клеток, получивших вектор со вставкой.
разрезание и сшивание
Плазмидную ДНК обрабатывают теми же рестриктазами, и она превращается в линейную молекулу, если подобрана такая рестриктаза, которая вносит в плазмиду 1 разрыв. В результате на концах всех образующихся фрагментов ДНК оказываются одни и те же липкие концы. При понижении температуры эти концы соединяются случайным образом, и их сшивают ДНК-лигазой (см. рис. 3).

Получают смесь кольцевых ДНК разного состава: некоторые из них будут содержать определённую последовательность ДНК хромосомной ДНК, соединённую с бактериальной ДНК, другие – соединённые вместе фрагменты хромосомной ДНК, а третьи – восстановленную кольцевую плазмиду или её димер (Рис. 4).

трансформация
Далее этой смесью проводят генетическую трансформацию бактерий, не содержащих плазмиды. Трансформация - процесс поглощения клеткой организма свободной молекулы ДНК из среды и встраивания её в геном, что приводит к появлению у такой клетки новых для неё наследуемых признаков, характерных для организма-донора ДНК. В каждую клетку может проникнуть и размножиться там только одна плазмида. Такие клетки помещают на твёрдую питательную среду, в которой содержится антибиотик тетрациклин. Клетки, в которые не попала плазмида, на этой среде расти не будут, а клетки, несущие плазмиду, образуют колонии, в каждой из которых находятся потомки только одной клетки, т.е. все клетки в колонии несут одну и ту же плазмиду (см. рис. 5).

Селекция
Далее стоит задача выделить только клетки, в которые попал вектор со вставкой, и отличить их от клеток, несущих только вектор без вставки или вовсе не несущих вектора. Этот процесс отбора нужных клеток называется селекцией . Для этого применяют селективные маркеры - обычно гены устойчивости к антибиотикам в составе вектора, и селективные среды , содержащие антибиотики или другие вещества, обеспечивающие селекцию.
В рассматриваемом нами примере клетки из колоний, выросших в присутствии ампицилина, пересевают на две среды: в первой есть ампицилин, а во второй – тетрациклин. Колонии, содержащие только плазмиду, вырастут на обеих средах, а колонии, в плазмидах которых находится встроенная хромосомная ДНК на среде с тетрациклином не вырастут (рис. 5). Среди них специальными методами отбирают те, которые содержат интересующий нас ген, выращивают в достаточных количествах и выделяют плазмидную ДНК. Из неё с помощью тех же рестриктаз, которые использовались при получении рекомбинантной ДНК, вырезают интересующий индивидуальный ген. ДНК этого гена может использоваться для определения последовательности нуклеотидов, введения в какой либо организм для получения новых свойств или синтеза нужного белка. Такой метод выделения генов называется молекулярным клонированием .
ФЛУОРЕСЦЕНТНЫЕ БЕЛКИ
В качестве генов-маркеров при исследованиях эукариотических организмов очень удобно использовать флуоресцентные белки. Ген первого флуоресцентного белка, зеленого флуоресцирующего белка (green fluorescent protein, GFP) был выделен из медузы Aqeuorea victoria и внедрен в различные модельные организмы (см. рис. 6) В 2008 году О. Симомура, М. Чалфи и Р. Тсьен получили Нобелевскую премию за открытие и применение этого белка.

Затем были выделены гены других флуоресцентных белков - красного, синего, желтого. Эти гены были модифицированы искусственно, чтобы получить белки с нужными свойствами. Разнообразие флуоресцентных белков показано на рис. 7, где изображена чашка Петри с бактериями, содержащими гены различных флуоресцентных белков.

применение флуоресцентных белков
Ген флуоресцентного белка можно сшивать с геном любого другого белка, тогда при трансляции будет образовываться единый белок - трансляционно слитый белок, или фьюжн (fusion protein), который флуоресцирует. Таким образом можно изучать, например, локализацию (расположение) любых интересующих белков в клетке, их перемещение. При помощи экспрессии флуоресцентных белков только в определенных типах клеток можно помечать клетки этих типов в многоклеточном организме (см. рис. 8 - мозг мыши, в котором отдельные нейроны имеют разные цвета за счет определенной комбинации генов флуоресцентных белков). Флуоресцентные белки - незаменимый инструмент современной молекулярной биологии.

ПЦР
Еще один метод получения генов называется полимеразной цепной реакцией (ПЦР) . В его основе лежит способность ДНК-полимераз достраивать вторую нить ДНК по комплементарной нити, как это происходит в клетках при репликации ДНК.
Точки начала репликации в этом методе задаются двумя небольшими фрагментами ДНК, называемыми затравками,
или праймерами
. Эти затравки комплементарны концам интересующего гена на двух цепях ДНК. Сначала хромосомную ДНК, из которой надо выделить ген, смешивают с затравками и нагревают до 99 о С. Это приводит к разрыву водородных связей и расхождению нитей ДНК. После этого температуру понижают до50-70 о С (в зависимости от длины и последовательности затравок). В этих условиях затравки присоединяются к комплементарным участкам хромосомной ДНК, образуя правильную двойную спираль (см. рис. 9). После этого добавляют смесь всех четырёх нуклеотидов, нужных для синтеза ДНК, и ДНК-полимеразу. Фермент удлиняет затравки, строя двуспиральную ДНК от места прикрепления затравок, т.е. от концов гена, до конца одноцепочечной хромосомной молекулы.
Если теперь снова нагреть смесь, то хромосомные и вновь синтезированные цепи разойдутся. После охлаждения к ним снова присоединятся затравки, которые берутся в большом избытке (см. рис. 10).

На вновь синтезированных цепях они присоединятся не к тому концу, с которого начинался первый синтез а к противоположному, так как цепи ДНК антипараллельны. Поэтому во втором цикле синтеза на таких цепях достроится только последовательность, соответствующая гену (см. рис. 11).
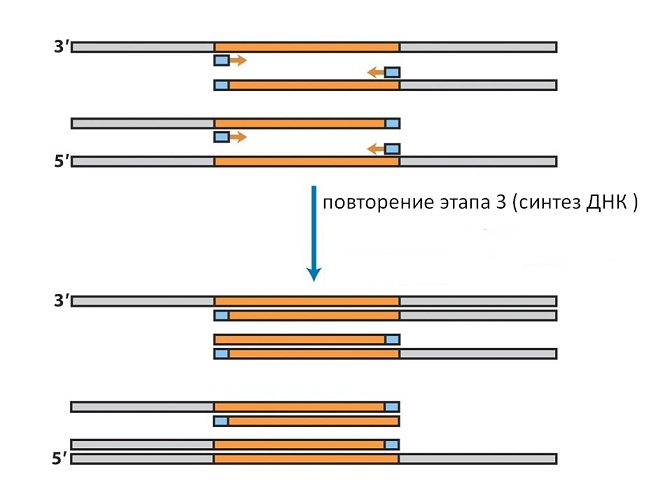
В данном методе применяется ДНК-полимераза из термофильных бактерий, способная выдерживать кипячение и работающая при температурах 70-80 о С, её не надо добавлять каждый раз, а достаточно внести в начале опыта. Повторяя процедуры нагрева и охлаждения в той же последовательности, мы можем в каждом цикле удваивать число последовательностей, ограниченных с двух концов внесёнными затравками (см. рис. 12).

После примерно 25 таких циклов число копий гена увеличится более чем в миллион раз. Такие количества легко можно отделить от внесённой в пробирку хромосомной ДНК и использовать для различных целей.
секвенирование ДНК
Ещё одним важным достижением является разработка методов определения последовательности нуклеотидов в ДНК - секвенирования ДНК (от англ. sequence - последовательность). Для этого необходимо получить чистые от других ДНК гены одним из описанных методов. Затем цепи ДНК разделяют нагреванием и прибавляют к ним затравку, меченую радиоактивным фосфором или флуоресцентной меткой. Обратите внимание, что берётся одна затравка, комплементарная одной цепи. Затем добавляется ДНК полимераза и смесь из 4-х нуклеотидов. Такая смесь делится на 4 части и к каждой добавляется один из нуклеотидов, модифицированный так, что у третьего атома дезоксирибозы он не содержит гидроксильной группы. Если такой нуклеотид включится в синтезируемую цепь ДНК, то её удлинение не сможет продолжаться, т.к. полимеразе некуда будет присоединять следующий нуклеотид. Поэтому синтез ДНК после включения такого нуклеотида обрывается. Таких нуклеотидов, называемых дидезоксинуклеотиды, добавляется значительно меньше, чем обычных, поэтому обрыв цепи происходит лишь изредка и в каждой цепи в разных местах. В результате получается смесь цепей разной длины, на конце каждой из них стоит один и тот же нуклеотид. Таким образом длина цепи соответствует номеру нуклеотида в изучаемой последовательности, например, если у нас был адениловый дидезоксинуклеотид, а полученные цепи имели длину 2, 7 и 12 нуклеотидов, значит в гене во второй, седьмой и двенадцатой позиции стоял аденин. Полученную смесь цепей легко разделить по размерам при помощи электрофореза, а синтезированные цепи выявить по радиоактивности на рентгеновской плёнке (см. рис. 10).

Получается картина, приведённая внизу рисунка, называемая радиоавтографом. Двигаясь по нему снизу вверх и читая буква над колонками каждой зоны мы получим последовательность нуклеотидов, приведённую на рисунке справа от автографа. Оказалось, что синтез останавливается не толоко дидезоксинуклеотидами, но и нуклеотидами, у которых в третьем положении сахара присоединяется какая-нибудь химическая группа, например флюоресцентный краситель. Если каждый нуклеотид пометить своим красителем, то зоны, получаемые при разделении синтезированных цепей, будут светиться разным светом. Это позволяет проводить реакцию в одной пробирке одновременно для всех нуклеотидов и разделяя полученные цепи по длине, идентифицировать нуклеотиды по цвету (см. рис. 11).

Такие методы позволили определить последовательности не только отдельных генов, но и прочитать целые геномы. В настоящее время разработаны ещё более быстрые методы определения последовательностей нуклеотидов в генах. Если парвый геном человека был расшифрован большим международным консорциумом с использованием первого приведённого метода за 12 лет, второй, с использованием второго, за три года, то сейчас это может быть сделано за месяц. Это позволяет предсказывать предрасположенность человека к многим заболеваниям и заранее принимать меры, чтобы избежать их.
Можно сказать, что молекулярная биология исследует проявления жизни на неживых структурах или системах с элементарными признаками жизнедеятельности (которыми могут быть отдельные биологические макромолекулы, их комплексы или органеллы), изучая, каким образом ключевые процессы, характеризующие живую материю, реализуются посредством химических взаимодействий и превращений.
Выделение молекулярной биологии из биохимии в самостоятельную область науки продиктовано тем, что её главной задачей является изучение структуры и свойств биологических макромолекул, участвующих в различных процессах, выяснение механизмов их взаимодействия. Биохимия же занимается изучением собственно процессов жизнедеятельности, закономерностей их протекания в живом организме и превращений молекул, сопровождающих эти процессы. В конечном счёте, молекулярная биология пытается ответить на вопрос, зачем происходит тот или иной процесс, тогда как биохимия отвечает на вопросы где и как с точки зрения химии происходит рассматриваемый процесс.
История
Молекулярная биология как отдельное направление биохимии начала формироваться в 30-х годах прошлого столетия. Именно тогда для углублённого понимания феномена жизни возникла необходимость в целенаправленных исследованиях на молекулярном уровне процессов хранения и передачи наследственной информации в живых организмах. Тогда и определилась задача молекулярной биологии в изучении структуры, свойств и взаимодействия нуклеиновых кислот и белков. Термин «молекулярная биология» был впервые употреблен английским учёным Уильямом Астбери в контексте исследований, касавшихся выяснения зависимостей между молекулярной структурой и физическими и биологическими свойствами фибриллярных белков, таких, как коллаген, фибрин крови или сократительные белки мышц.
На заре возникновения молекулярной биологии РНК считалась компонентом растений и грибов, а ДНК рассматривалась как типичный компонент животных клеток. Первым исследователем, доказавшим, что ДНК содержится в растениях, был Андрей Николаевич Белозерский , выделивший в 1935 году ДНК гороха. Это открытие установило тот факт, что ДНК является универсальныой нуклеиновой кислотой, присутствующей в клетках растений и животных.
Серьёзным достижением стало установление Джорджем Бидлом и Эдуардом Татумом прямой причинно-следственной связи между генами и белками. В своих экспериментах они подвергали клетки нейроспоры (Neurospora crassa ) ретгеновскому облучению, вызывавшему мутации. Полученные результаты показали, что это приводило к изменению свойств специфических ферментов.
В 1940 году Альбер Клод выделил из цитоплазмы животных клеток цитоплазматические РНК-содержащие гранулы, которые были меньше митохондрий. Он назвал их микросомами. Впоследствии при исследовании структруы и свойств выделенных частиц была установлена их основополагающая роль в процессе биосинтеза белка. В 1958 году на первом симпозиуме, посвящённом этим частицам, было принято решение называть эти частицы рибосомами.
Ещё одним важным шагом в развитии молекулярной биологии стали опубликованные в 1944 г. данные эксперимента Освальда Эвери, Колина МакЛауда и Маклина МакКарти, показавшие, что причиной трансформации бактерий является ДНК. Это было первое экспериментальное доказательство роли ДНК в передаче наследственной информации, развенчавшее бытовавшее ранее представление о белковой природе генов.
В начале 50-х годов Фредерик Сэнгер показал, что белковая цепь является уникальной последовательностью аминокислотных остатков. В конце 50-х годов Макс Перуц и Джон Кендрю расшифровали пространственное строение первых белков. Уже в 2000 году были известны сотни тысяч природных аминокислотных последовательностей и тысячи пространственных структур белков.
Примерно в то же время исследования Эрвина Чаргаффа позволили ему сформулировать правила, описывающие соотношение азотистых оснований в ДНК (правила гласят, что независимо от видовых различий в ДНК количество гуанина равно количеству цитозина, а количество аденина равно количеству темина), что помогло в дальнейшем сделать величайший прорыв в молекулярной биологии и одно из величайших открытий в биологии вообще.
Это событие произошло в 1953 году, когда Джеймс Уотсон и Фрэнсис Крик , основываясь на работах Розалинды Франклин и Мориса Уилкинса по рентгено-структурному анализу ДНК, установили двухспиральную структуру молекулы ДНК. Это открытие позволило ответить на принципиальный вопрос о способности носителя наследственной информации к самовоспроизведению и понять механизм передачи такой информации. Этими же учеными был сформулирован принцип комплементарности азотистых оснований, имеющий ключевое значение для понимания механизма образования надмолекулярных структур. Это принцип, применяемый теперь для описания всех молекулярных комплексов, позволяет описывать и предсказывать условия возникновения слабых (невалентных) межмолекулярных взаимодействий, обуславливающих возможность формирования вторичной, третичной и т.д. структуры макромолекул, протекания самосборки надмолекулярных биологических систем, определяющих столь большое разнообразие молекулярных структур и их функциональных наборов. Тогда же, в 1953 году возник научный журнал Journal of Molecular Biology. Его возглавил Джон Кендрю, сферой научных интересов которого было исследование структуры глобулярных белков (Нобелевская премия 1962 года совместно с Максом Перуцем). Аналогичный русскоязычный журнал под названием «Молекулярная биология» был основан в СССР В. А. Энгельгардтом в 1966 году.
В 1958 году Фрэнсис Крик сформулировал т.н. центральнаю догму молекулярной биологии: представление о необратимости потока генетической информации от ДНК через РНК к белкам по схеме ДНК→ДНК (репликация, создание копии ДНК), ДНК→РНК (транскрипция , копирование генов), РНК→ белок (трансляция, декодирование информации о структуре белков). Эта догма в 1970 году была несколько поправлена с учётом накопленных знаний, поскольку было открыто явление обратной транскрипции независимо Ховардом Темином и Дэвидом Балтимором: был обнаружен фермент - ревертаза, отвечающий за осуществление обратной транскрипции - образования двуцепочечной ДНК на матрице одноцепочечной РНК, которое происходит у онкогенных вирусов. Следует отметить, что строгая необходимость потока генетической информации от нуклеиновых кислот к белкам до сих пор остаётся основой молекулярной биологии.
В 1957 году Александр Сергеевич Спирин совместно с Андреем Николаевичем Белозерским показали, что, при существенных различиях в нуклеотидном составе ДНК из разных организмов, состав суммарных РНК сходен. На основании этих данных они пришли к сенсационному заключению о том, что суммарная РНК клетки не может выступать в качестве переносчика генетической информации от ДНК к белкам, поскольку не соответствует ей по своему составу. Вместе с тем они заметили, что существует минорная фракция РНК, которая полностью соответствует по своему нуклеотидному составу ДНК и которая может быть истинным переносчиком генетической инфрмации от ДНК к белкам. В результате они предсказали существование относительно небольших молекул РНК, являющихся по строению аналогами отдельных участков ДНК и выполняющих роль посредников при передаче генетической информации, содержащейся в ДНК, в рибосому, где с использованием этой информации осуществляется синтез белковых молекул. В 1961 году (С. Бреннер , Ф. Жакоб , М. Месельсон с одной стороны и Ф. Гро, Франсуа Жакоб и Жак Моно первыми получили опытное подтверждение существования таких молекул - информационной (матричной) РНК. Тогда же они разработали концепцию и модель функциональной единицы ДНК - оперона, которая позволила объяснить, как именно осуществляется регуляция экспрессии генов у прокариот. Исследование механизмов биосинтеза белка и принципов структурной организации и работы молекулярных машин - рибосом - позволило сформулировать постулат, описывающий движение генетической информации, называемый центральной догмой молекулярной биологии: ДНК - иРНК - белок.
В 1961 году и в течение последующих нескольких лет Хайнрихом Маттэем и Маршаллом Ниренбергом, а затем Харом Кораной и Робертом Холли были проведены несколько работ по расшифровке генетического кода, в результате которых была установлена непосредственная взаимосвязь между структурой ДНК и синтезируемыми белками и определена последовательность нуклеотидов, определяющая набор аминокислот в белке. Также были получены данные об универсальности генетического кода. Открытия были отмечены нобелевской премией 1968 года.
Для развития современных представлений о функциях РНК решающим было открытие некодирующих РНК, сделанное по результатам работ Александра Сергеевича Спирина совместно с Андреем Николаевичем Белозерским 1958 года, Чарльзом Бреннером с соавторами и Солом Шпигельманом 1961 года. Этот вид РНК составляют основную часть клеточных РНК. К некодирующим в первую очередь относятся рибосомные РНК.
Серьёзное развитие получили способы культивирования и гибридизации животных клеток. В 1963 году Франсуа Жакобом и Сиднеема Бреннером были сформулированы представления о репликоне - последовательности неотъемлемо реплицирующихся генов, объясняющей важные аспекты регуляции репликации генов.
В 1967 году в лаборатории А. С. Спирина было впервые продемонстрировано, что форма компактно свёрнутой РНК определяет морфологию рибосомной частицы.
В 1968 году было сделано значительное фундаментальное открытие. Оказаки, обнаружив фрагменты ДНК отстающей цепи при исследовании процесса репликации, названные в честь неё фрагментами Оказаки, уточнила механизм репликации ДНК.
В 1970 году независимо Ховардом Темином и Дэвидом Балтимором было сделано значительное открытие: был обнаружен фермент - ревертаза, отвечающий за осуществление обратной транскрипции - образования двуцепочечной ДНК на матрице одноцепочечной РНК, которое происходит у онкогенных вирусов, содержащих РНК.
Ещё одним важным достижением молекулярной биологии стало объяснение механизма мутаций на молекулярном уровне. В результате серии исследований были установлены основные типы мутаций: дупликации, инверсии, делеции, транслокации и транспозиции. Это дало возможность рассматривать эволюционные изменения с точки зрения генных процессов, позволило разработать теорию молекулярных часов, которая применяется в филогении.
К началу 70-х годов были сформулированы основные принципы функционирования нуклеиновых кислот и белков в живом организме. Было установлено, что белки и нуклеиновые кислоты в организме синтезируются по матричному механизму, молекула-матрица несёт в себе зашифрованную информацию о последовательности аминокислот (в белке) или нуклеотидов (в нуклеиновой кислоте). При репликации (удвоении ДНК) или транскрипции (синтезе иРНК) такой матрицей служит ДНК, при трансляции (синтезе белка) или обратной транскрипции - иРНК.
Таким образом, были созданы теоретические предпосылки для развития прикладных направлений молекулярной биологии, в частности, генетической инженерии . В 1972 году Пол Берг , Герберт Боер и Стэнли Коэн разработали технологию молекулярного клонирования. Тогда ими впервые была получена в пробирке рекомбинантная ДНК. Эти выдающиеся эксперименты заложили основы генетической инженерии, а этот год считается датой рождения этого научного направления.
В 1977 году Фредерик Сэнгер, и независимо Аллан Максам и Уолтер Гилберт разработали различные методы определения первичной структуры (секвенирования) ДНК. Метод Сэнгера, так называемый метод обрыва цепи, является основой современного метода секвенирования. Принцип секвенирования основан на использовании меченых оснований, выступающих в качестве терминаторов в циклической реакции секвенирования. Этот метод получил широкое распространение благодаря возможности быстро проводить анализ.
1976 г. - Фредерик. Сэнгер расшифровал нуклеотидную последовательность ДНК фага φΧ174 длиной 5375 нуклеотидных пар.
1981 г. - серповидноклеточная анемия становится первой генетической болезнью, диагностируемой с помощью анализа ДНК.
1982-1983 открытие каталитической функции РНК в американских лабораториях Т. Чека и С. Олтмана изменило существовавшее представления об исключительной роли белков. По аналогии с каталитическими белками - энзимами, каталитические РНК были названы рибозимами.
1987 год Кери Мюллез открыл полимеразную цепную реакцию, благодаря которой возможно искусственно значительно увеличить количество молекул ДНК в растворе для дальнейшей работы. На сегодняшний день это один из наиболее важных методов молекулярной биологии, применяющийся при исследовании наследственных и вирусных заболеваний, при изучении генов и при генетическом установлении личности и установлении родства и т.п.
В 1990 году одновременно тремя группами учёных был опубликован метод, позволявший быстро получать в лаборатории синтетические функционально активные РНК (искусственные рибозимы или молекулы, взаимодействующие с различными лигандами - аптамеры). Этот метод получил название «эволюция в пробирке». А вскоре после этого, в 1991-1993 года в лаборатории А.Б. Четверина была экспериментально показана возможность существования, роста и амплификации молекул РНК в форме колоний на твёрдых средах.
В 1998 году практически одновременно Крейг Мелло и Эндрю Фаер описали наблюдавшийся ранее при генных экспериментах с бактериями и цветами механизм РНК-интерференции , при котором небольшая двухцепочечная молекула РНК приводит к специфичному подавлению экспрессии гена.
Открытие механизма РНК-интерференции имеет очень важное практическое значение для современной молекулярной биологии. Это явление широко используется в научных экспериментах в качестве инструмента для «выключения», то есть, подавления экспрессии отдельных генов. Особый интерес вызван тем, что этот способ позволяет осуществлять обратимое (временное) подавление активности изучаемых генов. Ведутся исследования возможности применения этого явления для лечения вирусных, опухолевых, дегенеративных и метаболических заболеваний. Следует отметить, что в 2002 году были открыты мутанты вирусы полиомиелита, способные избегать РНК-интерференции, поэтому требуется ещё кропотливая работа для разработки действительно эффективных методов лечения на основе этого явления.
В 1999-2001 годах несколькими группами исследователей определена с разрешением от 5,5 до 2,4 ангстрем структура бактериальной рибосомы.
Предмет
Достижения молекулярной биологии в познании живой природы трудно переоценить. Больших успехов удалось достичь благодаря удачной концепции исследований: сложные биологические процессы рассматриваются с позиции отдельных молекулярных систем, что позволяет применять точные физико-химические методы исследования. Это также привлекло в эту область науки много великих умов из смежных направлений: химии, физики, цитологии, вирусологии , что также благотворно повлияло на масштабы и скорость развития научных знаний в этой области. Столь значимые открытия, как определение структуры ДНК, расшифровка генетического кода, искусственная направленная модификация генома, позволили значительно глубже понять специфику процессов развития организмов и успешно решать многочисленные важнейшие фундаментальные и прикладные научные, медицинские и социальные задачи, которые ещё не так давно считались неразрешимыми.
Предметом изучения молекулярной биологии являются в основном белки, нуклеиновые кислоты и молекулярные комплексы (молекулярные машины) на их основе и процессы, в которых они участвуют.
Нуклеиновые кислоты представляют собой линейные полимеры, состоящие из нуклеотидных звеньев (соединений пятичленного сахара с фосфатной группой при пятом атоме цикла и одного из четырёх азотистых оснований), соединённых между собой сложноэфирной связью фосфатных групп. Таким образом, нуклеиновая кислота - это пентозофосфатный полимер с азотистыми основаниями в качестве боковых заместителей. Химический состав цепочки РНК отличается от ДНК тем, что первая состоит из пятичленного цикла углевода рибозы, тогда как вторая - из дегидрокслилированного производного рибозы - дезоксирибозы. При этом пространственно эти молекулы различаются кардинально, поскольку РНК - это гибкая одноцепочечная молекула, тогда как ДНК - это двуцепочечная молекула.
Белки - это линейные полимеры, представляющие собой цепочки альфа-аминокислот, соединённых между собой пептидной связью, откуда их второе название - полипептиды. В состав природных белков входит множество различных аминокислотных звеньев - у человека до 20 -, что определяет широкое разнообразие функциональных свойств этих молекул. Те или иные белки принимают участие почти в каждом процессе в организме и выполняют множество задач: играют роль клеточного строительного материала, обеспечивают транспорт веществ и ионов, катализируют химические реакции, - список этот очень длинный. Белки образуют устойчивые молекулярные конформации различного уровня организации (вторичные и третичные структуры) и молекулярные комплексы, что ещё больше расширяет их функционал. Эти молекулы могут обладать высокой специфичностью к выполнению каких-либо задач благодаря образованию сложной пространственной глобулярной структуры. Большое разнообразие белков обеспечивает постоянный интерес учёных к этому виду молекул.
Современные представления о предмете молекулярной биологии основаны на обобщении, выдвинутом впервые в 1958 году Фрэнсисом Криком как центральная догма молекулярной биологии. Суть её заключалась в утверждении, что генетическая информация в живых организмах проходит строго определённые этапы реализации: копирование из ДНК в ДНК входе наследования, из ДНК в РНК, а затем из РНК в белок, причём обратный переход не осуществим. Это утверждение было справедливо лишь от части, поэтому впоследствии центральная догма была поправлена с оглядкой на открывшиеся новые данные.
На данный момент известно несколько путей реализации генетического материала, представляющих различные последовательности осуществления трёх видов существования генетической информации: ДНК, РНК и белок. В девяти возможных путях реализации выделяют три группы: это три общих превращения (general), осуществляющиеся в норме в большинстве живых организмов; три особых превращения(special), осуществляющиеся в некоторых вирусах или в особых лабораторных условиях; три неизвестных превращения (unknown), осуществление которых, как считается, невозможно.
К общим превращениям относятся следующие пути реализации генетического кода: ДНК→ДНК (репликация), ДНК→РНК (транскрипция), РНК→белок (трансляция).
Для осуществления передачи наследственных признаков родителям необходимо передать потомкам полноценную молекулу ДНК. Процесс, благодаря которому на основе исходной ДНК может быть синтезирована её точная копия, а следовательно, может быть передан генетический материал, называется репликацией. Он осуществляется специальными белками, которые распутывают молекулу (выпрямляют её участок), расплетают двойную спираль и при помощи ДНК-полимеразы создают точную копию исходной молекулы ДНК.
Для обеспечения жизнедеятельности клетки ей необходимо постоянно обращаться к генетическому коду, заложенному в двойной спирали ДНК. Однако эта молекула слишком велика и неповоротлива для применения её в качестве непосредственного источника генетического материала для непрерывного синтеза белка. Поэтому в ходе реализации информации, заложенной в ДНК, есть посредническая стадия: синтез иРНК, представляющей собой небольшую одноцепочечную молекулу, комплементарную определённому отрезку ДНК, кодирующему некоторый белок. Процесс транскрипции обеспечивается РНК-полимеразой и факторами транскрипции. Полученная молекула затем может быть легко доставлена в отдел клетки, ответственный за синтез белка - рибосому.
После попадания и РНК в рибосому наступает заключительная стадия реализации генетической информации. При этом рибосома считывает с иРНК генетический код триплетами , называющимися кодонами и синтезирует на основе получаемой информации соответствующий белок.
В ходе особых превращений генетический код реализуется по схеме РНК→РНК (репликация), РНК→ДНК (обратная транскрипция), ДНК→белок (прямая трансляция). Репликация такого вида реализуется во многих вирусах, где она осуществляется ферментом РНК-зависимой РНК-полимеразой. Аналогичные ферменты находятся и в клетках эукариот, где они связаны с процессом РНК-глушения (silencing). Обратная транскрипция обнаружена в ретровирусах, где она осуществляется под действием фермента обратной транскриптазы, а также в некоторых случаях в эукариотических клетках, например, при теломерном синтезе. Прямая трансляция осуществляется только в искусственных условиях в изолированной системе вне клетки.
Любой из трех возможных переходов генетической информации из белка в белок, РНК или ДНК считается невозможным. Случай воздействия прионов на белки, в результате которого образуется аналогичный прион, условно можно было бы отнести к виду реализации генетической информации белок→белок. Тем не менее, формально он таковым не является, поскольку не затрагивает аминокислотную последовательность в белке.
Любопытна история возникновения термина «центральная догма». Поскольку слово догма в общем случае означает утверждение, которое не подлежит сомнению, а само слово имеет явный религиозный подтекст, выбор его в качестве описания научного факта не совсем правомерен. По признанию самого Фрэнсиса Крика, это была его ошибка. Он хотел придать выдвигаемой теории большей значимости, выделить её на фоне остальных теорий и гипотез; для чего решил использовать это величественное, по его представлению, слово, не понимая его истинного смысла. Название это, однако, прижилось.
Молекулярная биология сегодня
Бурное развитие молекулярной биологии, постоянный интерес к достижениям в этой области со стороны общества и объективная важность исследований привели к возникновению большого числа крупных научно-исследовательских центров молекулярной биологии по всему миру. Среди крупнейших следует упомянуть следующие: лаборатория молекулярной биологии в Кембридже, Королевский институт в Лондоне - в Великобритании; институты молекулярной биологии в Париже, Марселе и Страсбурге, Пастеровский институт - во Франции; отделы молекулярной биологии в Гарвардском университете и Массачусетском технологическом институте, университете в Беркли, в Калифорнийском технологическом институте, в Рокфеллеровском университете, в институте здравоохранения в Бетесде - в США; институты Макса Планка, университеты в Гёттингене и Мюнхене, Центральный институт молекулярной биологии в Берлине, институты в Йене и Халле - в Германии; Каролинский институт в Стокгольме в Швеции.
В России ведущими центрами в этой области являются Институт молекулярной биологии им. В.А.Энгельгардта РАН, Институт молекулярной генетики РАН, Институт биологии гена РАН, Институт физико-химической биологии им. А. Н. Белозерского МГУ им. М.В.Ломоносова, Институт биохимии им. А.Н.Баха РАН и Институт белка РАН в Пущино.
Сегодня область интересов молекулярных биологов охватывает широкий спектр фундаментальных научных вопросов. По-прежнему ведущую роль занимает изучение структуры нуклеиновых кислот и биосинтеза белка, исследования строения и функций различных внутриклеточных структур, и клеточных поверхностей. Также важными направления исследований являются изучение механизмов рецепции и передачи сигналов, молекулярных механизмов транспорта соединений внутри клетки а также из клетки во внешнюю среду и обратно. Cреди основных направлений научного поиска в области прикладной молекулярной биологии одной из наиболее приоритетных является проблема возникновения и развития опухолей. Также очень важным направлением, изучением которого занимается раздел молекулярной биологии - молекулярная генетика, является изучение молекулярных основ возникновения наследственных заболеваний, и вирусных заболеваний, например, СПИДа, а также разработка способов их предупреждения и, возможно, лечения на генном уровне. Широкое применение нашли открытия и разработки молекулярных биологов в судебной медицине. Настоящая революция в области идентификации личности была сделана в 80-х годах учёными из России, США и Великобритании благодаря разработке и внедрению в повседневную практику метода «геномной дактилоскопии» - установления личности по ДНК. Исследования в этой области не прекращаются и по сей день, современные методы позволяют устанавливать личность с вероятностью ошибки одна миллиардная процента. Уже сейчас идёт активная разработка проекта генетического паспорта, что, как предполагается, позволит сильно снизить уровень преступности.
Методология
Сегодня молекулярная биология располагает обширным арсеналом методов, позволяющих решать самые передовые и самые сложные задачи, стоящие перед учёными.
Одним из самых распространённых методов в молекулярной биологии является гель-электрофорез , который решает задачи разделения смеси макромолекул по размеру или по заряду. Почти всегда после разделения макромолекул в геле применяется блоттинг , метод, позволяющий переносить макромолекулы из геля (сорбировать ) на поверхность мембраны для удобства дальнейшей работы с ними, в частности гибридизации . Гибридизация - формирование гибридной ДНК из двух цепей, имеющих различную природу, - метод, играющий важную роль в фундаментальных исследованиях. Он применяется для определения комплементарных отрезков в разных ДНК (ДНК разных видов), с его помощью происходит поиск новых генов, с его помощью была открыта РНК интерференция, а его принцип лёг в основу геномной дактилоскопии.
Большую роль в современной практике молекулярно-биологических исследований играет метод секвенирования - определения последовательности нуклеотидов в нуклеиновых кислотах и аминокислот в белках.
Современную молекулярную биологию невозможно представить без метода полимеразной цепной реакции (ПЦР). Благодаря этому методу осуществляется увеличение количества (амплификация) копий некоторой последовательности ДНК, чтобы позволяет получить из одной молекулы достаточное количество вещества для дальнейшей работы с ним. Аналогичный результат достигается технологией молекулярного клонирования, в которой требующаяся нуклеотидная последовательность внедряется в ДНК бактерии (живых систем), после чего размножение бактерий приводит к необходимому результату. Этот подход технически значительно сложнее, однако позволяет одновременно получать результат экспрессии исследуемой нуклеотидной последовательности.
Также в молекулярно-биологических исследованиях широко применяются методы ультрацентрифугирование (для разделения макромолекул (больших количеств), клеток, органелл), методы электронной и флуоресцентной микроскопии, спектрофотометрические методы, рентгеноструктурный анализ, авторадиография, и т.п.
Благодаря техническому прогрессу и научным изысканиям в области химии, физики, биологии и информатики современное оборудование позволяет выделять, изучать и изменять отдельные гены и процессы, в которые они вовлечены.
1. Введение.
Предмет, задачи и методы молекулярной биологии и генетики. Значение "классической" генетики и генетики микроорганизмов в становлении молекулярной биологии и генной инженерии. Понятие гена в "классической" и молекулярной генетике, его эволюция. Вклад методологии генной инженерии в развитие молекулярной генетики. Прикладное значение генной инженерии для биотехнологии.
2. Молекулярные основы наследственности.
Понятие о клетке, ее макромолекулярный состав. Природа генетического материала. История доказательства генетической функции ДНК.
2.1. Различные виды нуклеиновых кислот. Биологические функции нуклеиновых кислот. Химическое строение, пространственная структура и физические свойства нуклеиновых кислот. Особенности строения генетического материала про - и эукариот. Комплементарные пары оснований Уотсона-Крика. Генетический код. История расшифровки генетического кода. Основные свойства кода: триплетность, код без запятых, вырожденность. Особенности кодового словаря, семьи кодонов, смысловые и «бессмысленные» кодоны. Кольцевые молекулы ДНК и понятие о сверхспирализации ДНК. Топоизомеры ДНК и их типы. Механизмы действия топоизомераз. ДНК-гираза бактерий.
2.2. Транскрипция ДНК. РНК-полимераза прокариот, ее субъединичная и трехмерная структуры. Разнообразие сигма-факторов. Промотор генов прокариот, его структурные элементы. Стадии транскрипционного цикла. Инициация, образование “открытого комплекса”, элонгация и терминация транскрипции. Аттенюация транскрипции. Регуляция экспрессии триптофанового оперона. “Рибопереключатели”. Механизмы терминации транскрипции. Негативная и позитивная регуляция транскрипции. Лактозный оперон. Регуляция транскрипции в развитии фага лямбда. Принципы узнавания ДНК регуляторными белками (САР-белок и репрессор фага лямбда). Особенности транскрипции у эукариот. Процессинг РНК у эукариот. Кепирование, сплайсинг и полиаденилирование транскриптов. Механизмы сплайсинга. Роль малых ядерных РНК и белковых факторов. Альтернативный сплайсинг, примеры.
2.3. Трансляция , ее этапы, функция рибосом. Локализация рибосом в клетке. Прокариотический и эукариотический типы рибосом; 70S и 80S рибосомы. Морфология рибосом. Подразделение на субчастицы (субъединицы). Кодон-зависимое связывание аминоацил-тРНК в элонгационном цикле. Кодон-антикодоновое взаимодействие. Участие фактора элонгации EF1 (EF-Tu) в связывании аминоацил-тРНК с рибосомой. Фактор элонгации EF1В (EF-Ts), его функция, последовательность реакций с его участием. Антибиотики, воздействующие на этап кодон-зависимого связывания аминоацил-тРНК с рибосомой. Аминогликозидые антибиотики (стрептомицин, неомицин, канамицин, гентамицин и др.), механизм их действия. Тетрациклины как ингибиторы связывания аминоацил-тРНК с рибосомой. Инициация трансляции. Основные этапы процесса инициации. Инициация трансляции у прокариот: факторы инициации, инициаторные кодоны, 3¢-конец РНК малой рибосомной субчастицы и последовательность Шайна-Дальгарно в мРНК. Инициация трансляции у эукариот: факторы инициации, инициаторные кодоны, 5¢-нетранслируемая область и кэп-зависимая «концевая» инициация. «Внутренняя» кэп-независимая инициация у эукариот. Транспептидация. Ингибиторы транспептидации: хлорамфеникол, линкомицин, амицетин, стрептограмины, анизомицин. Транслокация. Участие фактора элонгации EF2 (EF-G) и ГТФ. Ингибиторы транслокации: фусидовая кислота, виомицин, их механизмы действия. Терминация трансляции. Терминирующие кодоны. Белковые факторы терминации прокариот и эукариот; два класса факторов терминации и механизмы их действия. Регуляция трансляции у прокариот.
2.4. Репликация ДНК и ее генетический контроль. Полимеразы, участвующие в репликации, характеристика их ферментативных активностей. Точность воспроизведения ДНК. Роль стерических взаимодействий между парами оснований ДНК при репликации. Полимеразы I, II и III E. coli. Субъединицы полимеразы III. Вилка репликации, “ведущая” и “отстающая” нити при репликации. Фрагменты Оказаки. Комплекс белков в репликационной вилке. Регуляция инициации репликации у E. соli. Терминация репликации у бактерий. Особенности регуляции репликации плазмид. Двунаправленная репликация и репликация по типу катящегося кольца.
2.5. Рекомбинация , ее типы и модели. Общая или гомологичная рекомбинация. Двухнитевые разрывы ДНК, инициирующие рекомбинацию. Роль рекомбинации в пострепликативной репарации двухнитевых разрывов. Структура Холлидея в модели рекомбинации. Энзимология общей рекомбинации у E. coli. RecBCD комплекс. RecA белок. Роль pекомбинации в обеспечении синтеза ДНК при повреждениях ДНК, прерывающих репликацию. Рекомбинация у эукариот. Ферменты рекомбинации у эукариот. Сайт-специфичная рекомбинация. Различия молекулярных механизмов общей и сайт-специфичной рекомбинации. Классификация рекомбиназ. Типы хромосомных перестроек, осуществляемых при сайт-специфичной рекомбинации. Регуляторная роль сайт-специфичной рекомбинации у бактерий. Конструирование хромосом многоклеточных эукариот с помощью системы сайт-специфичной рекомбинации фага.
2.6. Репарация ДНК. Классификация типов репарации. Прямая репарация тиминовых димеров и метилированного гуанина. Вырезание оснований. Гликозилазы. Механизм репарации неспаренных нуклеотидов (mismatch репарация). Выбор репарируемой нити ДНК. SOS-репарация. Свойства ДНК полимераз, участвующих в SOS-репарации у прокариот и эукариот. Представление об “адаптивных мутациях” у бактерий. Репарация двухнитевых разрывов: гомологичная пострепликативная рекомбинация и объединение негомологичных концов молекулы ДНК. Взаимосвязь процессов репликации, рекомбинации и репарации.
3. Мутационный процесс.
Роль биохимических мутантов в формировании теории один ген – один фермент. Классификация мутаций. Точковые мутации и хромосомные перестройки, механизм их образования. Спонтанный и индуцированный мутагенез. Классификация мутагенов. Молекулярный механизм мутагенеза. Взаимосвязь мутагенеза и репарации. Идентификация и селекция мутантов. Супрессия: внутригенная, межгенная и фенотипическая.
4. Внехромосомные генетические элементы.
Плазмиды, их строение и классификация. Половой фактор F, его строение и жизненный цикл. Роль фактора F в мобилизации хромосомного переноса. Образование доноров типа Hfr и F". Механизм конъюгации. Бактериофаги, их структура и жизненный цикл. Вирулентные и умеренные бактериофаги. Лизогения и трансдукция. Общая и специфическая трансдукция. Мигрирующие генетические элементы: транспозоны и IS-последовательности, их роль в генетическом обмене. ДНК-транспозоны в геномах прокариот и эукариот. IS-последовательности бактерий, их структура. IS-последовательности как компонент F-фактора бактерий, определяющего способность передачи генетического материала при конъюгации. Транспозоны бактерий и эукариотических организмов. Прямой нерепликативный и репликативный механизмы транспозиций. Представление о горизонтальном переносе транспозонов и их роли в структурных перерстройках (эктопическая рекомбинация) и в эволюции генома.
5. Исследование структуры и функции гена.
Элементы генетического анализа. Цис-транс комплементационный тест. Генетическое картирование с использованием конъюгации, трансдукции и трансформации. Построение генетических карт. Тонкое генетическое картирование. Физический анализ структуры гена. Гетеродуплексный анализ. Рестрикционный анализ. Методы секвенирования. Полимеразная цепная реакция. Выявление функции гена.
6. Регуляция экспрессии генов. Концепции оперона и регулона. Контроль на уровне инициации транскрипции. Промотор, оператор и регуляторные белки. Позитивный и негативный контроль экспрессии генов. Контроль на уровне терминации транскрипции. Катаболит-контролируемые опероны: модели лактозного, галактозного, арабинозного и мальтозного оперонов. Аттенюатор-контролируемые опероны: модель триптофанового оперона. Мультивалентная регуляция экспрессии генов. Глобальные системы регуляции. Регуляторный ответ на стрессы. Посттранскрипционный контроль. Сигальная трансдукция. Регуляция с участием РНК: малые РНК, сенсорные РНК.
7. Основы генной инженерии. Ферменты рестрикции и модификации. Выделение и клонирование генов. Векторы для молекулярного клонирования. Принципы конструирования рекомбинантных ДНК и их введения в реципиентные клетки. Прикладные аспекты генной инженерии.
а). Основная литература:
1. Уотсон Дж., Туз Дж., Рекомбинантные ДНК: Краткий курс. – М.: Мир, 1986.
2. Гены. – М.: Мир. 1987.
3. Молекулярная биология: структура и биосинтез нуклеиновых кислот. / Под ред. . – М. Высшая шк. 1990.
4. , – Молекулярная биотехнология. М. 2002.
5. Спирин рибосомы и биосинтез белка. – М.: Высшая школа, 1986.
б). Дополнительная литература:
1. Хесин генома. – М.: Наука. 1984.
2. Рыбчин генетической инженерии. – СПб.: СПбГТУ. 1999.
3. Патрушев генов. – М.: Наука, 2000.
4. Современная микробиология. Прокариоты (в 2-х тт.). – М.: Мир, 2005.
5. М. Сингер, П. Берг. Гены и геномы. – М.: Мир, 1998.
6. Щелкунов инженерия. – Новосибирск: Из-во Сиб. Унив., 2004.
7. Степанов биология. Структура и функции белков. – М.: В. Ш., 1996.
Развитие биохимии, биофизики, генетики, цитохимии, многих разделов микробиологии и вирусологии примерно к началу 40-х годов XX в. вплотную подвело к изучению жизненных явлений на молекулярном уровне. Успехи, достигнутые этими науками, одновременно и с разных сторон привели к осознанию того факта, что именно на молекулярном уровне функционируют основные управляющие системы организма и что дальнейший прогресс этих наук будет зависеть от раскрытия биологических функций молекул, составляющих тела организмов, их участия в синтезе и распаде, взаимных превращениях и репродукции соединений в клетке, а также происходящего при этом обмена энергией и информацией. Так на стыке этих биологических дисциплин с химией и физикой возникла совершенно новая отрасль - молекулярная биология.
В отличие от биохимии, внимание современной молекулярной биологии сосредоточено преимущественно на изучении структуры и функции важнейших классов биополимеров - белков и нуклеиновых кислот, первые из которых определяют самую возможность протекания обменных реакций, а вторые - биосинтез специфических белков. Понятно поэтому, что провести четкое разграничение молекулярной биологии и биохимии, соответствующих разделов генетики, микробиологии и вирусологии невозможно.
Возникновение молекулярной биологии было тесно связано с разработкой новых методов исследования, о которых уже говорилось в соответствующих главах. Наряду с развитием электронной микроскопии и других методов микроскопической техники большую роль сыграли разработанные в 50-х годах методы фракционирования клеточных элементов. Они основывались на усовершенствованных методах дифференциального центрифугирования (А. Клод, 1954). К этому времени уже имелись довольно надежные методы выделения и фракционирования биополимеров. Сюда относится, в частности, предложенный А. Тизелиусом (1937; Нобелевская премия, 1948) метод фракционирования белков при помощи электрофореза, методы выделения и очистки нуклеиновых кислот (Е. Кей, А. Даунс, М. Севаг, А. Мирский и др.). Параллельно во многих лабораториях мира разрабатывались различные методы хроматографического анализа (А. Мартин и Р. Синг, 1941; Нобелевская премия, 1952), впоследствии существенно усовершенствованные.
Неоценимую услугу в расшифровке структуры биополимеров сыграл рентгеноструктурный анализ. Основные принципы рентгеноструктурного анализа были разработаны в Королевском колледже Лондонского университета под руководством У. Брегга группой исследователей, куда входили Дж. Бернал, А. Лондсдейл, У. Астбери, Дж. Робертсон и др.
Следует особо отметить исследования профессора Московского государственного университета А. Р. Кизеля по биохимии протоплазмы (1925 - 1929), имевшие важнейшее значение для последующего становления молекулярной биологии. Кизель нанес удар прочно укоренившемуся представлению, что в основе всякой протоплазмы лежит особое белковое тело - пластин, будто бы определяющее все ее важнейшие структурные и функциональные особенности. Он показал, что пластин - это белок, который встречается только у миксомицетов, и то на определенной стадии развития, и что никакого постоянного компонента - единого скелетного белка - в протоплазме не существует. Тем самым изучение проблемы строения протоплазмы и функциональной роли белков вышло на правильный путь и получило простор для своего развития. Исследования Кизеля завоевали мировое признание, стимулировав изучение химии составных частей клетки.
Термин "молекулярная биология", впервые использованный английским кристаллографом профессором Лидского университета У. Астбери, появился, вероятно, в начале 40-х годов (до 1945 г.). Основополагающие рентгеноструктурные исследования белков и ДНК, проведенные Астбери в 30-х годах, послужили основой для последующей успешной расшифровки вторичной структуры этих биополимеров. В 1963 г. Дж. Бернал писал: "Памятник ему будет установлен всей молекулярной биологией - наукой, которую он назвал и действительно основал" * , В литературе этот термин появился впервые, пожалуй, в 1946 г. в статье У. Астбери "Прогресс рентгеноструктурного анализа органических и фибриллярных соединений", опубликованной в английском журнале "Природа" ** . В своей Гарвеевской лекции Астбери (1950) отмечал: "Мне приятно, что сейчас термин молекулярная биология уже довольно широко употребляется, хотя мало вероятно, что я первым предложил его. Он мне нравился и я уже давно старался его распространять" *** . Уже в 1950 г. Астбери было ясно, что молекулярная биология имеет дело прежде всего со структурой и конформацией макромолекул, изучение которых имеет решающее значение для понимания функционирования живых организмов.
* (Biogr. Mem. Fellows Roy. Soc, 1963, v. 9, 29. )
** (W. T. Astbury. Progress of X-ray analysis of organic and fibre structures.- Nature,. 1946, v. 157, 121. )
*** (W. T. Astbury. Adventures in Molecular Biology. Thomas Springfield, 1952, p. 3. )
Перед молекулярной биологией стояли и стоят, собственно, те же задачи, что и перед всей биологией в целом,- познание сущности жизни и ее основных явлений, в частности таких, как наследственность и изменчивость. Современная молекулярная биология в первую очередь призвана расшифровать структуру и функцию генов, пути и механизмы реализации генетической информации организмов на разных стадиях онтогенеза и на разных этапах ее считывания. Она призвана вскрыть тонкие механизмы регуляции активности генов и клеточной дифференцировки, выяснить природу мутагенеза и молекулярные основы эволюционного процесса.
Установление генетической роли нуклеиновых кислот
Для становления молекулярной биологии наибольшее значение имели следующие открытия. В 1944 г. американские исследователи О. Эвери, К. Мак-Леод (Нобелевская премия, 1923) и М. Мак-Карти показали, что выделенные из пневмококков молекулы ДНК обладают трансформирующей активностью. После гидролиза этих ДНК дезоксирибонуклеазой их трансформирующая активность полностью исчезала. Тем самым впервые было убедительно доказано, что генетическими функциями в клетке наделена именно ДНК, а не белок.
Справедливости ради следует заметить, что явление бактериальной трансформации было обнаружено значительно ранее открытия Эвери, Мак-Леода и Мак-Карти. В 1928 г. Ф. Гриффит опубликовал статью, в которой сообщил, что после добавления к невирулентным (некапсулированным) пневмококкам убитых клеток капсулированного вирулентного штамма получаемая смесь клеток становится губительной для мышей. Более того, выделяемые из зараженных этой смесью животных живые клетки пневмококков были уже вирулентными и обладали полисахаридной капсулой. Тем самым в этом опыте было показано, что под воздействием каких-то компонентов убитых клеток пневмококков некапсулированная форма бактерий превращается в капсулообразующую вирулентную форму. 16 лет спустя Эвери, Мак-Леод и Мак-Карти заменили в этом опыте убитые целые клетки пневмококков их дезоксирибонуклеиновой кислотой и показали, что именно ДНК обладает трансформирующей активностью (см. также главы 7 и 25). Значение этого открытия трудно переоценить. Оно стимулировало изучение нуклеиновых кислот во многих лабораториях мира и заставило сконцентрировать внимание ученых именно на ДНК.
Наряду с открытием Эвери, Мак-Леода и Мак-Карти к началу 50-х годов уже накопилось довольно большое количество прямых и косвенных данных о том, что нуклеиновые кислоты играют исключительную роль в жизнедеятельности и несут генетическую функцию. На это, в частности, указывал и характер локализации ДНК в клетке и данные Р. Вендрели (1948) о том, что содержание ДНК на клетку строго постоянно и коррелирует со степенью плоидности: в гаплоидных половых клетках ДНК вдвое меньше, чем в диплоидных соматических. В пользу генетической роли ДНК свидетельствовала также ее выраженная метаболическая стабильность. К началу 50-х годов накопилось много разнообразных фактов, свидетельствовавших о том, что большинство известных мутагенных факторов действуют преимущественно на нуклеиновые кислоты и, в особенности, на ДНК (Р. Хочкисс, 1949; Г. Эфрусси-Тейлор, 1951; Э. Фриз, 1957 и др.).
Особое значение в установлении генетической роли нуклеиновых кислот имело изучение различных фагов и вирусов. В 1933 г. Д. Шлезингер нашел ДНК в бактериофаге кишечной палочки. С момента выделения У. Стенли (1935, Нобелевская премия, 1946) вируса табачной мозаики (ВТМ) в кристаллическом состоянии начался новый этап в изучении растительных вирусов. В 1937 - 1938 гг. сотрудники Ротамстедской сельскохозяйственной станции (Англия) Ф. Боуден и Н. Пири показали, что многие выделенные ими растительные вирусы являются не глобулинами, а представляют собой рибонуклеопротеиды и содержат в качестве обязательного компонента нуклеиновую кислоту. В самом начале 40-х годов были опубликованы работы Г. Шрамма (1940), П. А. Агатова (1941), Г. Миллера и У. Стенли (1941), свидетельствовавшие о том, что заметная химическая модификация белкового компонента не приводит к утрате инфекционности ВТМ. Это указывало на то, что белковый компонент не может быть носителем наследственных свойств вируса, как продолжали считать многие микробиологи. Убедительные доказательства в пользу генетической роли нуклеиновой кислоты (РНК) у растительных вирусов были получены в 1956 г. Г. Шраммом в Тюбингене (ФРГ) и X. Френкель-Конратом в Калифорнии (США). Эти исследователи практически одновременно и независимо друг от друга выделили из ВТМ РНК и показали, что именно она, а не белок, обладает инфекционностью: в результате заражения растений табака этой РНК в них происходило формирование и размножение нормальных вирусных частиц. Это означало, что РНК содержит информацию для синтеза и сборки всех вирусных компонентов, в том числе и вирусного белка. В 1968 г. И. Г. Атабеков установил, что белок играет существенную роль при самом заражении растений - природой белка определяется спектр растений-хозяев.
В 1957 г. Френкель-Конрат впервые осуществил реконструкцию ВТМ из составляющих его компонентов - РНК и белка. Наряду с нормальными частицами он получил смешанные "гибриды", у которых РНК была от одного штамма, а белок - от другого. Наследственность таких гибридов полностью определялась РНК, и потомство вирусов принадлежало к тому штамму, РНК которого была использована для получения исходных смешанных частиц. Позднее опыты А. Гирера, Г. Шустера и Г. Шрамма (1958) и Г. Витмана (1960 - 1966) показали, что химическая модификация нуклеинового компонента ВТМ приводит к появлению разнообразных мутантов этого вируса.
В 1970 г. Д. Балтимор и Г. Темин установили, что перенос генетической информации может происходить не только от ДНК к РНК, но и наоборот. Они обнаружили у некоторых онкогенных РНК-содержащих вирусов (онкорнавирусы) особый фермент, так называемую обратную транскриптазу, который способен на цепях РНК комплементарно синтезировать ДНК. Это крупное открытие позволило понять механизм встройки в геном хозяина генетической информации РНК-содержащих вирусов и по-новому взглянуть на природу их онкогенного действия.
Открытие нуклеиновых кислот и изучение их свойств
Термин нуклеиновые кислоты был введен немецким биохимиком Р. Альтманом в 1889 г., после того как эти соединения были открыты в 1869 г. швейцарским врачом Ф. Мишером. Мишер экстрагировал клетки гноя разбавленной соляной кислотой в течение нескольких недель и получил в остатке почти чистый ядерный материал. Этот материал он считал характерным" веществом клеточных ядер и назвал его нуклеином. По своим свойствам нуклеин резко отличался от белков: он был более кислым, не содержал серу, но зато в нем было много фосфора, он хорошо растворялся в щелочах, но не растворялся в разбавленных кислотах.
Результаты своих наблюдений над нуклеином Мишер направил Ф. Гоппе-Зейлеру для опубликования в журнале. Описанное им вещество было настолько необычным (в то время из всех биологических фосфорсодержащих соединений был известен только лецитин), что Гоппе-Зейлер не поверил опытам Мишера, вернул ему рукопись и поручил своим сотрудникам Н. Плошу и Н. Любавину проверить его выводы на другом материале. Работа Мишера "О химическом составе клеток гноя" вышла в свет двумя годами позже (1871). В то же время были опубликованы работы Гоппе-Зейлера и его сотрудников о составе клеток гноя, эритроцитов птиц, змей и других клеток. В течение последующих трех лет нуклеин был выделен из животных клеток и дрожжей.
В своей работе Мишер отмечал, что детальное изучение разных нуклеинов может привести к установлению различий между ними, предвосхитив тем самым идею специфичности нуклеиновых кислот. Исследуя молоки лосося, Мишер установил, что нуклеин находится в них в виде соли и связан с основным белком, который он назвал протамином.
В 1879 г. в лаборатории Гоппе-Зейлера изучением нуклеинов начал заниматься А. Коссель. В 1881 г. он выделил из нуклеина гипоксантин, однако в то время он еще сомневался в происхождении этого основания и считал, что гипоксантин может быть продуктом деградации белков. В 1891 г. в числе продуктов гидролиза нуклеина Коссель обнаружил аденин, гуанин, фосфорную кислоту и еще одно вещество со свойствами сахара. За исследования по химии нуклеиновых кислот Косселю в 1910 г. была присуждена Нобелевская премия.
Дальнейшие успехи в расшифровке структуры нуклеиновых кислот связаны с исследованиями П. Левина и сотрудников (1911 - 1934). В 1911 г. П. Левин и В. Жакобс идентифицировали углеводный компонент аденозина и гуанозина; они установили, что в состав этих нуклеозидов входит D-рибоза. В 1930 г. Левин показал, что углеводным компонентом дезоксирибонуклеозидов является 2-дезокси-D-рибоза. Из его работ стало известно, что нуклеиновые кислоты построены из нуклеотидов, т. е. фосфорилированных нуклеозидов. Левин считал, что основным типом связи в нуклеиновых кислотах (РНК) является 2", 5"-фосфодиэфирная связь. Это представление оказалось ошибочным. Благодаря работам английского химика А. Тодда (Нобелевская премия, 1957) и его сотрудников, а также английских биохимиков Р. Маркхема и Дж. Смита в начале 50-х годов стало известно, что основным типом связи в РНК является 3", 5"-фосфодиэфирная связь.
Левин показал, что разные нуклеиновые кислоты могут различаться по природе углеводного компонента: одни из них содержат сахар дезоксирибозу, а другие - рибозу. Кроме того, указанные два типа нуклеиновых кислот различались по природе одного из оснований: в нуклеиновых кислотах пентозного типа содержался урацил, а в нуклеиновых кислотах дезоксипентозного типа - тимин. Дезоксипентозную нуклеиновую кислоту (по современной терминологии, дезоксирибонуклеиновая кислота - ДНК) обычно легко выделяли в больших количествах из тимуса (зобной железы) телят. Поэтому она получила название тимонуклеиновой кислоты. Источником нуклеиновой кислоты пентозного типа (РНК) служили главным образом дрожжи и зародыши пшеницы. Этот тип часто называли дрожжевой нуклеиновой кислотой.
В начале 30-х годов довольно прочно укоренилось представление, будто для растительных клеток характерна нуклеиновая кислота дрожжевого типа, а тимонуклеиновая кислота свойственна только ядрам животных клеток. Два типа нуклеиновых кислот - РНК и ДНК - в то время называли соответственно растительной и животной нуклеиновыми кислотами. Однако, как показали ранние исследования А. Н. Белозерского, такое деление нуклеиновых кислот неоправданно. В 1934 г. Белозерский впервые обнаружил тимонуклеиновую кислоту в растительных клетках: из проростков гороха он выделил и идентифицировал тимин-пиримидиновое основание, характерное именно для ДНК. Затем он обнаружил тимин и в других растениях (семенах сои, фасоли). В 1936 г. А. Н. Белозерский и И. И. Дубровская выделили препаративно ДНК из проростков конского каштана. Кроме того, серия работ, выполненных в 40-х годах в Англии Д. Девидсоном с сотрудниками, убедительно показала, что растительная нуклеиновая кислота (РНК) содержится во многих животных клетках.
Широкое применение разработанной Р. Фельгеном и Г. Розенбеком (1924) цитохимической реакции на ДНК и реакции Ж. Браше (1944) на РНК позволило довольно быстро и однозначно решить вопрос о преимущественной локализации этих нуклеиновых кислот в клетке. Оказалось, что ДНК сосредоточена в ядре, в то время как РНК преимущественно концентрируется в цитоплазме. Позднее было выяснено, что РНК содержится как в цитоплазме, так и в ядре, а кроме того, были выявлены цитоплазматические ДНК.
Что касается вопроса о первичной структуре нуклеиновых кислот, то к середине 40-х годов в науке прочно утвердилось представление П. Левина, согласно которому все нуклеиновые кислоты построены по одному типу и состоят из одинаковых так называемых тетрануклеотидных блоков. В каждом из этих блоков, по мнению Левина, содержится четыре разных нуклеотида. Тетрануклеотидная теория строения нуклеиновых кислот в значительной мере лишала эти биополимеры специфичности. Поэтому не удивительно, что всю специфику живого связывали в то время только с белками, природа мономеров которых гораздо разнообразнее (20 аминокислот).
Первую брешь в теории тетрануклеотидного строения нуклеиновых кислот пробили аналитические данные английского химика Дж. Гуланда (1945 - 1947). При определении состава нуклеиновых кислот по азоту оснований он не получил эквимолярного соотношения оснований, как это должно было бы быть согласно теории Левина. Окончательно тетрануклеотидная теория строения нуклеиновых кислот рухнула в результате исследований Э. Чаргаффа и его сотрудников (1949 - 1951). Для разделения оснований, выщепляющихся из ДНК в результате ее кислотного гидролиза, Чаргафф использовал хроматографию на бумаге. Каждое из этих оснований было точно определено спектрофотометрически. Чаргафф заметил значительные отклонения от эквимолярного соотношения оснований в ДНК разного происхождения и впервые определенно заявил, что ДНК обладает выраженной видовой специфичностью. Тем самым было покончено с гегемонией концепции о специфичности белка в живой клетке. Анализируя ДНК разного происхождения, Чаргафф открыл и сформулировал уникальные закономерности состава ДНК, вошедшие в науку под названием правил Чаргаффа. Согласно этим правилам, во всех ДНК, независимо от происхождения, количество аденина равно количеству тимина (А = Т), количество гуанина равно количеству цитозина (Г = Ц), количество пуринов равно количеству пиримидинов (Г + А = Ц + Т), количество оснований с 6-аминогруппами равно количеству оснований с 6-кетогруп-пами (А+Ц=Г+Т). Вместе с тем, несмотря на такие строгие количественные соответствия, ДНК разных видов отличаются по величине отношения А+Т:Г+Ц. В одних ДНК количество гуанина и цитозина преобладает над количеством аденина и тимина (эти ДНК Чаргафф назвал ДНК ГЦ-типа); другие ДНК содержали аденина и тимина больше, чем гуанина и цитозина (эти ДНК были названы ДНК АТ-типа). Полученные Чаргаффом данные по составу ДНК сыграли исключительную роль в молекулярной биологии. Именно они легли в основу открытия строения ДНК, сделанного в 1953 г. Дж. Уотсоном и Ф. Криком.
Еще в 1938 г. У. Астбери и Ф. Белл при помощи рентгеноструктурного анализа показали, что плоскости оснований в ДНК должны быть перпендикулярными к длинной оси молекулы и напоминать как бы стопку пластин, лежащих друг над другом. По мере совершенствования техники рентгеноструктурного анализа к 1952 - 1953 гг. накопились сведения, позволившие судить о длине отдельных связей и углах наклона. Это дало возможность с наибольшей вероятностью представить характер ориентации колец пентозных остатков в сахарофосфатном костяке молекулы ДНК. В 1952 г. С. Фарберг предложил две умозрительные модели ДНК, которые представляли сложенную или закрученную саму на себя однотяжную молекулу. Не менее спекулятивная модель строения ДНК была предложена в 1953 г. Л. Полингом (лауреат Нобелевской премии, 1954) и Р. Кори. В этой модели три закрученные цепи ДНК образовывали длинную спираль, стержень которой был представлен фосфатными группами, а основания располагались снаружи от него. К 1953 г. М. Уилкинс и Р. Франклин получили более четкие рентгеноструктурные картины ДНК. Их анализ показал полную несостоятельность моделей Фарберга, Полинга и Кори. Используя данные Чаргаффа, сопоставляя разные сочетания молекулярных моделей отдельных мономеров и данные рентгеноструктурного анализа, Дж. Уотсон и Ф. Крик в 1953 г. пришли к выводу, что молекула ДНК должна быть двутяжной спиралью. Правила Чаргаффа резко ограничили число возможных упорядоченных сочетаний оснований в предлагаемой модели ДНК; они подсказали Уотсону и Крику, что в молекуле ДНК должно быть специфическое спаривание оснований - аденина с тимином, а гуанина с цитозином. Иными словами аденину в одной цепи ДНК всегда строго соответствует тимин в другой цепи, а гуанину в одной цепи обязательно соответствует цитозин в другой. Тем самым Уотсон и Крик впервые сформулировали исключительной важности принцип комплементарного строения ДНК, согласно которому одна цепь ДНК дополняет другую, т. е. последовательность оснований одной цепи однозначно определяет последовательность оснований в другой (комплементарной) цепи. Стало очевидно, что уже в самой структуре ДНК заложена потенциальная возможность ее точного воспроизведения. Эта модель строения ДНК в настоящее время является общепризнанной. За расшифровку структуры ДНК Крику, Уотсону и Уилкинсу в 1962 г. была присуждена Нобелевская премия.
Следует отметить, что идея о механизме точного воспроизведения макромолекул и передаче наследственной информации зародилась в нашей стране. В 1927 г. Н, К. Кольцов высказал предположение, что при размножении клеток происходит репродукция молекул путем точного автокаталитического воспроизведения имевшихся материнских молекул. Правда, в то время Кольцов наделял этим свойством не молекулы ДНК, а молекулы белковой природы, о функциональном значении которых тогда ничего не было известно. Тем не менее сама мысль об автокаталитическом воспроизведении макромолекул и механизме передачи наследственных свойств оказалась пророческой: она стала руководящей идеей современной молекулярной биологии.
Проведенные в лаборатории А. Н. Белозерского А. С. Спириным, Г. Н. Зайцевой, Б. Ф. Ванюшиным, С. О. Урысон, А. С. Антоновым и другими многолетние исследования (1957-1974) состава ДНК у самых разнообразных организмов полностью подтвердили закономерности, обнаруженные Чаргаффом, и полное соответствие с молекулярной моделью строения ДНК, предложенной Уотсоном и Криком. Эти исследования показали, что ДНК разных бактерий, грибов, водорослей, актиномицетов, высших растений, беспозвоночных и позвоночных обладают специфичностью состава. Особенно резко различия в составе (содержании АТ-пар оснований) выражены у микроорганизмов, оказываясь важным таксономическим признаком. У высших растений и животных видовые вариации в составе ДНК выражены значительно слабее. Но это вовсе не означает, что ДНК у них менее специфична. Кроме состава оснований специфичность в большей степени определяется их последовательностью в цепях ДНК.
Наряду с обычными основаниями в составе ДНК и РНК были обнаружены дополнительные азотистые основания. Так, в составе ДНК растений и животных Г. Уайт (1950) нашел 5-метилцитозин, а Д. Данн и Дж. Смит (1958) обнаружили в некоторых ДНК метилированный аденин. Долгое время метилцитозин считался отличительной чертой генетического материала высших организмов. В 1968 г. А. Н. Белозерский, Б. Ф. Ванюшин и Н. А. Кокурина установили, что он может встречаться также и в ДНК бактерий.
В 1964 г. М. Голд и Дж. Хурвитц открыли новый класс ферментов, осуществляющих природную модификацию ДНК - ее метилирование. После этого открытия стало ясно, что минорные (содержащиеся в малых количествах) основания возникают уже на готовой полинуклеотидной цепи ДНК в результате специфического метилирования остатков цитозина и аденина в особых последовательностях. В частности, по данным Б. Ф. Ванюшина, Я. И. Бурьянова и А. Н. Белозерского (1969) метилирование аденина в ДНК кишечной палочки может происходить в терминирующих кодонах. По данным А. Н. Белозерского и сотрудников (1968 - 1970), а также М. Мезельсона (США) и В. Арбера (Швейцария) (1965 - 1969) метилирование придает молекулам ДНК уникальные индивидуальные черты и в сочетании с действием специфических нуклеаз является частью сложного механизма, который осуществляет контроль за синтезом ДНК в клетке. Иными словами, характер метилирования той или иной ДНК предопределяет вопрос о том, может ли она размножаться в данной клетке.
Практически в то же время началось выделение и интенсивное изучение ДНК-метилаз и рестрицирующих эндонуклеаз; в 1969 - 1975 гг. установлены нуклеотидные последовательности, узнаваемые в ДНК некоторыми из этих ферментов (X. Бойер, X. Смит, С. Линн, К. Муррей). При гидролизе разных ДНК рестрицирующим ферментом выщепляются довольно крупные фрагменты с одинаковыми "липкими" концами. Это дает возможность не только анализировать структуру генов, как это сделано у небольших вирусов (Д. Натанс, С. Адлер, 1973 - 1975), но и конструировать различные геномы. С открытием этих специфических ферментов рестрикции генетическая инженерия стала ощутимой реальностью. Встроенные в небольшие плазмидные ДНК гены различного происхождения уже легко вводят в различные клетки. Так, получен новый тип биологически активных плазмид, дающих устойчивость к некоторым антибиотикам (С. Коэн, 1973), введены рибосомальные гены лягушки и дрозофилы в плазмиды кишечной палочки (Дж. Морроу, 1974; X. Бойер, Д. Хогнесс, Р. Девис, 1974 - 1975). Таким образом, открыты реальные пути для получения принципиально новых организмов путем введения и встраивания в их генофонд разнообразных генов. Это открытие может быть направлено на благо всего человечества.
В 1952 г. Г. Уайт и С. Коэн обнаружили, что в ДНК Т-четных фагов содержится необычное основание - 5-оксиметилцитозин. Позднее из работ Е. Волькина и Р. Синсхеймера (1954) и Коэна (1956) стало известно, что остатки оксиметилцитозина могут быть полностью или частично глюкозидированы, в результате чего молекула фаговой ДНК оказывается защищенной от гидролитического действия нуклеаз.
В начале 50-х годов из работ Д. Данна и Дж. Смита (Англия), С. Заменхофа (США) и А. Вакера (ФРГ) стало известно, что в ДНК могут включаться многие искусственные аналоги оснований, замещая иногда до 50% тимина. Как правило, эти замещения приводят к ошибкам при репликации, транскрипции ДНК и трансляции и к появлению мутантов. Так, Дж. Мармур (1962) установил, что в ДНК некоторых фагов вместо тимина содержится оксиметилурацил. В 1963 г. И. Такахаши и Дж. Мармур обнаружили, что в ДНК одного из фагов вместо тимина содержится урацил. Таким образом, рухнул еще один принцип, по которому ранее разделяли нуклеиновые кислоты. Со времен работ П. Левина считалось, что отличительным признаком ДНК является тимин, а РНК - урацил. Стало ясно, что этот признак не всегда надежен, и принципиальным различием химической природы двух типов нуклеиновых кислот, как это представляется на сегодняшний день, служит только характер углеводного компонента.
При изучении фагов было вскрыто много необычных признаков организации нуклеиновых кислот. С 1953 г. считалось, что все ДНК представляют собой двутяжные линейные молекулы, а РНК - только однотяжные. Это положение существенно поколебалось в 1961 г., когда Р. Синсхеймер обнаружил, что ДНК фага φ X 174 представлена однотяжной кольцевой молекулой. Правда, затем выяснилось, что в такой форме эта ДНК существует только в вегетативной фаговой частице, а репликативная форма ДНК этого фага также двутяжная. Кроме того, весьма неожиданным оказалось, что РНК некоторых вирусов могут быть двутяжными. Этот новый тип макромолекулярной организации РНК был обнаружен в 1962 г. П. Гоматосом, И. Таммом и другими исследователями у некоторых вирусов животных и у вируса раневой опухоли растений. Недавно В. И. Агол и А. А. Богданов (1970) установили, что помимо линейных молекул РНК существуют также замкнутые или циклические молекулы. Циклическая двутяжная РНК выявлена ими, в частности, у вируса энцефаломиэлокардита. Благодаря работам X. Дево, Л. Тиноко, Т. И. Тихоненко, Э. И. Будовского и других (1960 - 1974) стали известны основные черты организации (укладки) генетического материала у бактериофагов.
В конце 50-х годов американский ученый П. Доти установил, что при нагревании происходит денатурация ДНК, сопровождающаяся разрывом водородных связей между парами оснований и расхождением комплементарных цепей. Этот процесс носит характер фазового перехода по типу "спираль-клубок" и напоминает плавление кристаллов. Поэтому процесс тепловой денатурации ДНК Доти назвал плавлением ДНК. При медленном охлаждении происходит ренатурацпя молекул, т. е воссоединение комплементарных половинок.
Принцип ренатурации в 1960 г. был использован Дж. Мармуром и К. Шильдкраутом для определения степени "гибридизуемости" ДНК разных микроорганизмов. Впоследствии Е. Болтон и Б. Мак-Карти усовершенствовали этот прием, предложив метод так называемых ДНК-агаровых колонок. Этот метод оказался незаменимым в изучении степени гомологии нуклеотидной последовательности разных ДНК и выяснении генетического родства разных организмов. Открытая Доти денатурация ДНК в сочетании с описанной Дж. Манделем и А. Херши * (1960) хроматографией на метилированном альбумине и центрифугированием в градиенте плотности (метод разработан в 1957 г. М. Мезельсоном, Ф. Сталем и Д. Виноградом) широко используется для разделения, выделения и анализа отдельных комплементарных цепей ДНК Так, например, В. Шибальски (США), используя эти приемы для разделения ДНК лямбда фага, показал в 1967 - 1969 гг., что генетически активными являются обе цепочки фага, а не одна, как это было принято считать (С. Спигельман, 1961). Следует отметить, что впервые идея о генетической значимости обеих цепочек ДНК лямбда фага была высказана в СССР С. Е. Бреслером (1961).
* (За работы по генетике бактерий и вирусов А. Херши совместно с М. Дельбрюком и С. Луриа были удостоены в 1969 г. Нобелевской премии. )
Для понимания организации и функциональной активности генома первостепенное значение имеет определение нуклеотидной последовательности ДНК. Поиски методов такого определения ведутся во многих лабораториях мира. В США М. Бир с сотрудниками с конца 50-х годов пытается установить последовательность ДНК при помощи электронной микроскопии, но пока безуспешно. В начале 50-х годов из первых работ Синсхеймера, Чаргаффа и других исследователей по ферментативной деградации ДНК стало известно, что разные нуклеотиды в молекуле ДНК распределены хотя и нехаотично, но неравномерно. По данным английского химика К. Бартона (1961), пиримидины (их более 70%) сосредоточены в основном в виде соответствующих блоков. А. Л. Мазин и Б. Ф. Ванюшин (1968 - 1969) установили, что разные ДНК обладают различной степенью сблоченности пиримидинов и что в ДНК животных организмов она заметно возрастает по мере перехода от низших к высшим. Таким образом, эволюция организмов отражена и в структуре их геномов. Именно поэтому для понимания эволюционного процесса в целом сравнительное изучение структуры нуклеиновых кислот приобретает особое значение. Анализ структуры биологически важных полимеров и, в первую очередь, ДНК крайне важен и для решения многих частных вопросов филогенетики и таксономии.
Интересно отметить, что английский физиолог Э. Ланкестер, изучавший гемоглобины моллюсков, ровно 100 лет назад предвосхитивший идеи молекулярной биологии, писал: "Химические различия разных видов и родов животных и растений имеют такое же важное значение для выяснения истории их происхождения, как и различия в их форме. Если бы мы могли четко устанавливать различия в молекулярной организации и функционировании организмов, мы смогли бы значительно лучше разобраться в происхождении и эволюции разных организмов, чем на основании морфологических наблюдений" * . Значимость биохимических исследований для систематики подчеркивал и В. Л. Комаров, который писал, что "в основе всех даже чисто морфологических признаков, на основании которых мы классифицируем и устанавливаем виды, лежат именно биохимические различия" ** .
* (Е. R. Lankester. Uber das Vorcommen von Haemoglobin in den Muskeln der Mollusken und die Verbreitung desselben in den lebendigen Organismen.- "Pfluger"s Archiv fur die gesammte Physiol., 1871, Bd 4, 319. )
** (В. Л. Комаров. Избранные соч., т. 1. М.-Л., Изд-во АН СССР, 1945, стр. 331. )
А. В. Благовещенский и С. Л. Иванов еще в 20-х годах предприняли первые в нашей стране шаги по выяснению некоторых вопросов эволюции и систематики организмов на основе сравнительного анализа их биохимического состава (см. гл. 2). Сравнительный анализ структуры белков и нуклеиновых кислот в настоящее время становится все более ощутимым подспорьем для систематиков (см. главу 21). Этот метод молекулярной биологии позволяет не только уточнить положение отдельных видов в системе, но и заставляет по-новому взглянуть на сами принципы классификации организмов, а иногда и пересмотреть всю систему в целом, как это случилось, например, с систематикой микроорганизмов. Несомненно, и в будущем анализ структуры генома будет занимать центральное место в хемосистематике организмов.
Огромное значение для становления молекулярной биологии имела расшифровка механизмов репликации ДНК и транскрипции (см главу 24).
Биосинтез белка
Важный сдвиг в решении проблемы биосинтеза белка связан с успехами в изучении нуклеиновых кислот. В 1941 г. Т. Касперсон (Швеция) и в 1942 г. Ж. Браше (Бельгия) обратили внимание на то, что в тканях с активным белковым синтезом содержится повышенное количество РНК. Они пришли к выводу, что рибонуклеиновые кислоты играют определяющую роль в синтезе белка. В 1953 г. Е. Гейл и Д. Фокс, как будто, получили прямые доказательства непосредственного участия РНК в биосинтезе белка: по их данным, рибонуклеаза существенно подавляла включение аминокислот в лизатах бактериальных клеток. Аналогичные данные были получены В. Олфри, М. Дели и А. Мирским (1953) на гомогенатах печени. Позднее Э. Гейл отказался от высказанной им правильной идеи о ведущей роли РНК в белковом синтезе, ошибочно считая, что активация белкового синтеза в бесклеточной системе происходила под влиянием какого-то другого вещества неизвестной природы. В 1954 г. П. Замечник, Д. Литлфилд, Р. Б. Хесин-Лурье и другие обнаружили, что наиболее активное включение аминокислот происходит в богатых РНК фракциях субклеточных частиц - микросом. П. Замечник и Э. Келлер (1953 - 1954) обнаружили, что включение аминокислот заметно усиливалось в присутствии надосадочной фракции в условиях регенерации АТФ. П. Сикевиц (1952) и М. Хогланд (1956) выделили из надосадочной жидкости белковую фракцию (рН 5 фракция), которая была ответственной за резкое стимулирование включения аминокислот в микросомах. Наряду с белками в надосадочной жидкости был обнаружен особый класс низкомолекулярных РНК, которые теперь называют транспортными РНК (тРНК). В 1958 г. Хогланд и Замечник, а также П. Берг, Р. Свит и Ф. Аллен и многие другие исследователи обнаружили, что для активации каждой аминокислоты необходим свой особый фермент, АТФ и специфическая тРНК. Стало ясно, что тРНК выполняют исключительно функцию адаптеров, т. е. приспособлений, которые находят на нуклеиновой матрице (иРНК) место соответствующей аминокислоте в формирующейся белковой молекуле. Эти исследования полностью подтвердили адапторную гипотезу Ф. Крика (1957), предусматривавшую существование в клетке полинуклеотидных адапторов, необходимых для правильного расположения аминокислотных остатков синтезирующегося белка на нуклеиновой матрице. Уже много позднее французский ученый Ф. Шапвиль (1962) в лаборатории Ф. Липмана (Нобелевская премия, 1953) в США весьма остроумно и однозначно показал, что местоположение аминокислоты в синтезирующейся белковой молекуле полностью определяется той специфической тРНК, к которой она присоединена. Адапторная гипотеза Крика была развита в работах Хогланда и Замечника.
К 1958 г. стали известны следующие основные этапы белкового синтеза: 1) активация аминокислоты специфическим ферментом из "рН 5 фракции" в присутствии АТФ с образованием аминоациладенилата; 2) присоединение активированной аминокислоты к специфической тРНК с высвобождением аденозинмонофосфата (АМФ); 3) связывание аминоацил-тРНК (тРНК, нагруженная аминокислотой) с микросомами и включение аминокислот в белок с высвобождением тРНК. Хогланд (1958) отметил, что на последнем этапе белкового синтеза необходим гуанозинтрифосфат (ГТФ).
Транспортные РНК и синтез гена
После обнаружения тРНК начались активные поиски их фракционирования и определения нуклеотидной последовательности. Наибольших успеходов добился американский биохимик Р. Холли. В 1965 г. он установил структуру аланиновой тРНК из дрожжей. При помощи рибонуклеаз (гуаниловая РНК-аза и панкреатическая РНК-аза) Холли разделил молекулу нуклеиновой кислоты на несколько фрагментов, определил в каждом из них по отдельности нуклеотидную последовательность и затем реконструировал последовательность всей молекулы аланиновой тРНК. Этот путь анализа нуклеотидной последовательности получил название блочного метода. Заслуга Холли состояла главным образом в том, что он научился разделять молекулу РНК не только на мелкие куски, как это делали многие и до него, но и на крупные фрагменты (четвертинки и половинки). Это и дало ему возможность правильно собрать отдельные маленькие куски воедино и тем самым воссоздать полную нуклеотидную последовательность всей молекулы тРНК (Нобелевская премия, 1968).
Этот прием сразу же был принят на вооружение во многих лабораториях мира. В течение последующих двух лет в СССР и за рубежом была расшифрована первичная структура сразу нескольких тРНК. А. А. Баев (1967) и сотрудники впервые установили последовательность нуклеотидов в дрожжевой валиновой тРНК. К настоящему времени изучено уже более десятка различных индивидуальных тРНК. Своеобразный рекорд в определении нуклеотидной последовательности установлен в Кембридже Ф. Сенгером и Г. Браунли. Эти исследователи разработали удивительно изящный метод разделения олигонуклеотидов и установили последовательность так называемой 5 S (рибосомной) РНК из клеток кишечной палочки (1968). Эта РНК состоит из 120 нуклеотидных остатков и в отличие от тРНК не содержит дополнительных минорных оснований, которые заметно облегчают анализ нуклеотидной последовательности, служа уникальными ориентирами отдельных фрагментов молекулы. В настоящее время благодаря использованию метода Сенгера и Браунли успешно продвигается работа по изучению последовательности длинных рибосомных РНК и некоторых вирусных РНК в лаборатории Ж. Эбеля (Франция) и других исследователей.
А. А. Баев и сотрудники (1967) обнаружили, что разрезанная пополам валиновая тРНК восстанавливает свою макромолекулярную структуру в растворе и, несмотря на дефект в первичной структуре, обладает функциональной активностью исходной (нативной) молекулы. Этот подход - реконструкция разрезанной макромолекулы после удаления определенных фрагментов - оказался весьма перспективным. Он широко используется сейчас для выяснения функциональной роли отдельных участков тех или иных тРНК.
В последние годы достигнут большое успех в получении кристаллических препаратов индивидуальных тРНК. Сейчас в нескольких лабораториях в США и Англии удалось закристаллизовать уже многие тРНК. Это позволило исследовать стуруктуру тРНК при помощи рентгеноструктурного анализа. В 1970 г. Р. Бок представил первые рентгенограммы и трехмерные модели нескольких тРНК, созданные им в Висконсинском университете. Эти модели помогают определить локализацию отдельных функционально активных участков в тРНК и понять основные принципы функционирования этих молекул.
Важнейшее значение для раскрытия механизма синтеза белка и решения проблемы специфичности этого процесса имела расшифровка природы генетического кода (см. главу 24), которую без преувеличения можно рассматривать как ведущее завоевание естествознания XX в.
Раскрытие Р. Холли первичной структуры тРНК дало толчок работам Г. Кораны * (США) по синтезу олигонуклеотидов и направило их на путь синтеза определенной биологической структуры - молекулы ДНК, кодирующей аланиновую тРНК. Сделанные Кораной почти 15 лет назад первые шаги по химическому синтезу коротких олигонуклеотидов завершились в 1970 г. впервые осуществленным синтезом гена. Корана и его сотрудники сначала из отдельных нуклеотидов синтезировали химическим путем короткие фрагменты длиной в 8-12 нуклеотидных остатков. Эти фрагменты с заданной нуклеотидной последовательностью образовывали спонтанно двутяжные комплементарные куски с перекрыванием в 4 - 5 нуклеотидов. Затем эти готовые куски в нужном порядке поочередно соединяли конец в конец при помощи фермента ДНК-лигазы. Таким образом, в отличие от репликации молекул ДНК, по А. Корнбергу ** (см. главу 24), Коране удалось заново создать молекулу естественной двутяжной ДНК по заранее намеченной программе в соответствии с последовательностью тРНК, описанной Холли. Аналогичным образом сейчас ведутся работы по синтезу других генов (М. Н. Колосов, 3. А. Шабарова, Д. Г. Кнорре, 1970 - 1975).
* (За исследования генетического кода Г. Коране и М. Ниренбергу была присуждена в 1968 г. Нобелевская премия. )
** (За открытие полимеразы и синтез ДНК А. Корнбергу, а за синтез РНК С. Очоа в 1959 г. была присуждена Нобелевская премия. )
Микросомы, рибосомы, трансляция
В середине 50-х годов считалось, что центром белкового синтеза в клетке являются микросомы. Термин микросомы был впервые введен в 1949 г. А. Клодом для обозначения фракции мелких гранул. Позднее выяснилось, что за белковый синтез ответственна не вся фракция микросом, состоящая из мембран и гранул, а только мелкие рибонуклеопротеидные частицы. Эти частицы в 1958 г. были названы Р. Робертсом рибосомами.
Классические исследования бактериальных рибосом были проведены А. Тисьером и Дж. Уотсоном в 1958 - 1959 гг. Бактериальные рибосомы оказались несколько мельче растительных и животных. Дж. Литлтон (1960), М. Кларк (1964) и Э. Н. Светайло (1966) показали, что рибосомы хлоропластов высших растений и митохондрий принадлежат к бактериальному типу. А. Тисьер и другие (1958) обнаружили, что рибосомы диссоциируют на две неравные субъединицы, содержащие по одной молекуле РНК. В конце 50-х годов считалось, что каждая молекула рибосомной РНК состоит из нескольких коротких фрагментов. Однако А. С. Спирин в 1960 г. впервые показал, что РНК в субчастицах представлены непрерывной молекулой. Д. Уоллер (1960), разделив рибосомные белки при помощи электрофореза в крахмальном геле, установил, что они весьма гетерогенны. Первое время многие сомневались в данных Уоллера, поскольку казалось, что белок рибосомы должен быть строго гомогенным, как, например, белок ВТМ. В настоящее время в результате исследований Д. Уоллера, Р. Траута, П. Трауба и других биохимиков стало известно, что в состав собственно рибосомных частиц входит более 50 совершенно различных по структуре белков. А. С. Спирину в 1963 г. удалось впервые развернуть рибосомные субчастицы и показать, что рибосомы представляют собой компактно скрученный рибонуклеопротеидный тяж, который в определенных условиях может развертываться. В 1967 - 1968 гг. М. Номура полностью реконструировал биологически активную субчастицу из рибосомной РНК и белка и даже получил такие рибосомы, в которых белок и РНК принадлежали разным микроорганизмам.
До сегодняшнего дня неясна роль рибосомной РНК. Предполагается, что она является той уникальной специфической матрицей, на которой при формировании рибосомной частицы находит строго определенное место каждый из многочисленных рибосомных белков (А. С. Спирин, 1968).
А. Рич (1962) обнаружил агрегаты из нескольких рибосом, соединенных между собой нитью иРНК. Эти комплексы были названы полисомами. Обнаружение полисом позволило Ричу и Уотсону (1963) высказать предположение, что синтез полипептидной цепи происходит на рибосоме, которая как бы продвигается по цепочке иРНК. По мере продвижения рибосомы по цепочке иРНК в частице совершается считывание информации и образование полипептидной цепи белка, а новые рибосомы поочередно присоединяются к высвобождающемуся прочитанному концу иРНК. Из данных Рича и Уотсона следовало, что значение полисом в клетке состоит в массовой продукции белка путем последовательного прочитывания матрицы сразу несколькими рибосомами.
В результате исследований М. Ниренберга, С. Очоа, Ф. Липмана, Г. Кораны и других в 1963 - 1970 гг. стало известно, что наряду с иРНК, рибосомами, АТФ и аминоацил-тРНК в процессе трансляции принимает участие большое количество разнообразных факторов, а сам процесс трансляции может быть условно разделен на три этапа - инициацию, собственно трансляцию и терминацию.
Инициация трансляции означает синтез первой пептидной связи в комплексе рибосома - матричный полинуклеотид - аминоацил-тРНК. Такой инициаторной активностью обладает не всякая аминоацил-тРНК, а формилметионил-тРНК. Это вещество было впервые выделено в 1964 г. Ф. Сенгером и К. Маркером. С. Бретчер и К. Маркер (1966) показали, что инициаторная функция формилметионил-тРНК обусловлена ее повышенным сродством к пептидильному центру рибосомы. Для начала трансляции крайне важны также некоторые белковые факторы инициации, которые были выделены в лабораториях С. Очоа, Ф. Гро и других иссследовательских центрах. После образования первой пептидной связи в рибосоме начинается собственно трансляция, т. е. последовательное присоединение аминоацильного остатка к С-концу полипептида. Многие детали процесса трансляции изучили К. Монро и Дж. Бишоп (Англия), И. Рыхлик и Ф. Шорм (ЧССР), Ф. Липман, М. Бретчер, В. Гилберт (США) и другие исследователи. В 1968 г. А. С. Спирин для объяснения механизма работы рибосомы предложил оригинальную гипотезу. Приводным механизмом, обеспечивающим все пространственные перемещения тРНК и иРНК во время трансляции, является периодическое размыкание и смыкание субчастиц рибосомы. Окончание трансляции закодировано в самой считываемой матрице, которая содержит терминирующие кодоны. Как показал С. Бреннер (1965 - 1967), такими кодонами являются триплеты УАА, УАГ и УГА. М. Капеччи (1967) выявил также специальные белковые факторы терминации. А. С. Спириным и Л. П. Гавриловой описан так называемый "неферментативный" синтез белка в рибосомах (1972 - 1975) без участия белковых факторов. Это открытие важно для понимания происхождения и эволюции биосинтеза белка.
Регуляция активности генов и белков
После проблемы специфичности белкового синтеза на первом месте в молекулярной биологии оказалась проблема регуляции синтеза белков, или, что то же самое, регуляции активности генов.
Функциональная неравнозначность клеток и связанные с ней репрессия и активация генов давно привлекали внимание генетиков, но до последнего времени реальный механизм контроля генной активности оставался неизвестным.
Первые попытки объяснить регуляторную активность генов были связаны с изучением гистонных белков. Еще супруги Стэдман * в начале 40-х годов XX в. высказывали мысль, что именно гистоны могут играть в этом явлении основную роль. В дальнейшем они получили первые четкие данные о различиях в химической природе гистонных белков. В настоящее время количество фактов, свидетельствующих в пользу этой гипотезы, с каждым годом все более возрастает.
* (Е. Stedman, E. Stedman. The basic proteins of cell nuclei.- Phylosoph. Trans. Roy. Soc. London, 1951, v. 235, 565 - 595. )
В то же время накапливается все большее число данных, говорящих о том, что регуляция генной активности - гораздо более сложный процесс, чем простое взаимодействие участков генов с молекулами гистонных белков. В 1960 - 1962 гг. в лаборатории Р. Б. Хесина-Лурье было выяснено, что гены фагов начинают считываться неодновременно: гены фага Т2 можно разделить на ранние, функционирование которых происходило в первые минуты заражения бактериальной клетки, и поздние, начинавшие синтезировать иРНК после завершения работы ранних генов.
В 1961 г. французские биохимики Ф. Жакоб и Ж. Моно предложили схему регуляции активности генов, которая сыграла исключительную роль в понимании регуляторных механизмов клетки вообще. Согласно схеме Жакоба и Моно, в ДНК кроме структурных (информационных) генов имеются еще гены-регуляторы и гены-операторы. Ген-регулятор кодирует синтез специфического вещества - репрессора, который может присоединяться как к индуктору, так и к гену-оператору. Ген-оператор сцеплен со структурными генами, а ген-регулятор находится на некотором отдалении от них. Если в среде нет индуктора, например, лактозы, то синтезируемый геном-регулятором репрессор связывается с геном-оператором и, блокируя его, выключает работу всего оперона (блок структурных генов вместе с управляющим ими оператором). Образования фермента в этих условиях не происходит. Если же в среде появляется индуктор (лактоза), то продукт гена-регулятора - репрессор - связывается с лактозой и снимает блок с гена-оператора. В этом случае становится возможной работа структурного гена, кодирующего синтез фермента, и фермент (лактоза) появляется в среде.
По мнению Жакоба и Моно, эта схема регуляции применима ко всем адаптивным ферментам и может иметь место как при репрессии, когда образование фермента подавляется избытком продукта реакции, так и при индукции, когда внесение субстрата вызывает синтез фермента. За исследования регуляции активности генов Жакоб и Моно были удостоены в 1965 г. Нобелевской премии.
Первоначально эта схема казалась слишком надуманной. Однако впоследствии выяснилось, что регуляция генов по этому принципу имеет место не только у бактерий, но и у других организмов.
Начиная с 1960 г. заметное место в молекулярной биологии занимают исследования организации генома и структуры хроматина у эукариотических организмов (Дж. Боннер, Р. Бриттен, В. Олфри, П. Уокер, Ю. С. Ченцов, И. Б. Збарский и др.) и по регуляции транскрипции (А. Мирский, Г. П. Георгиев, М. Бернстил, Д. Голл, Р. Цанев, Р. И. Салганик). Долгое время оставалась неизвестной и спорной природа репрессора. В 1968 г. М. Пташне (США) показал, что репрессором является белок. Он выделил его в лаборатории Дж. Уотсона и обнаружил, что репрессор, действительно, обладает сродством к индуктору (лактозе) и одновременно "узнает" ген-оператор лак-оперона и специфически связывается с ним.
В последние 5 - 7 лет получены данные о наличии еще одной управляющей ячейки генной активности - промоторе. Оказалось, что по соседству с операторным участком, к которому присоединяется продукт, синтезированный на гене-регуляторе - белковом веществе репрессора, имеется другой участок, который также следует отнести к членам регуляторной системы генной активности. К этому участку присоединяется белковая молекула фермента РНК-полимеразы. В промоторном участке должно произойти взаимное узнавание уникальной последовательности нуклеотидов в ДНК и специфической конфигурации белка РНК-полимеразы. От эффективности узнавания будет зависеть осуществление процесса считывания генетической информации с данной последовательностью генов оперона, примыкающего к промотору.
Кроме описанной Жакобом и Моно схемы, в клетке существуют и другие механизмы регуляции генов. Ф. Жакоб и С. Бреннер (1963) установили, что регуляция репликации бактериальной ДНК определенным образом контролируется клеточной мембраной. Опыты Жакоба (1954) по индукции разных профагов убедительно показали, что под влиянием различных мутагенных факторов в клетке лизогенных бактерий начинается избирательная репликация гена профага, а репликация генома хозяина блокируется. В 1970 г. Ф. Белл сообщил о том, что в цитоплазму из ядра могут переходить небольшие молекулы ДНК и уже там транскрибироваться.
Таким образом, регуляция активности генов может осуществляться на уровне репликации, транскрипции и трансляции.
Значительные успехи достигнуты в изучении регуляции не только синтеза ферментов, но и их активности. На явления регуляции активности ферментов в клетке указывали еще в 50-х годах А. Новик и Л. Сциллард. Г. Умбаргер (1956) установил, что в клетке существует весьма рациональный путь подавления активности фермента конечным продуктом цепи реакций по типу обратной связи. Как было установлено Ж. Моно, Ж. Шанже, Ф. Жакобом, А. Парди и другими исследователями (1956 - 1960), регуляция активности ферментов может осуществляться по аллостерическому принципу. Фермент или одна из его субъединиц, кроме сродства к субстрату, обладает сродством к одному из продуктов цепи реакций. Под влиянием такого продукта-сигнала фермент так изменяет свою конформацию, что утрачивает активность. В результате вся цепь ферментативных реакций выключается в самом начале. На существенную роль конформационных изменений белка в ферментативных реакциях, а в известном смысле и на наличие аллостерического эффекта, указывали Д. Уимен и Р. Вудворд (1952; лауреат Нобелевской премии, 1965).
Структура и функции белков
В результате работ Т. Осборна, Г. Гофмейстера, А. Гюрбера, Ф. Шульца и многих других в конце XIX в. были получены многие животные и растительные белки в кристаллическом виде. Примерно в это же время при помощи разных физических методов были установлены молекулярные веса некоторых белков. Так, в 1891 г. А. Сабанеев и Н. Александров сообщили, что молекулярный вес овальбумина составляет 14 000; в 1905 г. Э. Рейд установил, что молекулярный вес гемоглобина равен 48 000. Полимерная структура белков была раскрыта в 1871 г. Г. Глазиветцом и Д. Габерманом. Идея о пептидной связи отдельных аминокислотных остатков в белках была высказана Т. Куртиусом (1883). Работы по химической конденсации аминокислот (Э. Шаал, 1871; Г. Шифф, 1897; Л. Бальбиано и Д. Траскиатти, 1900) и синтезу гетерополипептидов (Э. Фишер, 1902 - 1907, Нобелевская премия, 1902) привели к разработке основных принципов химической структуры белков.
Первый кристаллический фермент (уреаза) был получен в 1926 г. Дж. Самнером (Нобелевская премия, 1946), а в 1930 г. Дж. Нортроп (Нобелевская премия, 1946) получил кристаллический пепсин. После этих работ стало ясно, что ферменты имеют белковую природу. В 1940 г. М. Куниц выделил кристаллическую РНК-азу. К 1958 г. уже было известно более 100 кристаллических ферментов и свыше 500 ферментов, выделенных в некристаллическом виде. Получение высокоочищенных препаратов индивидуальных белков способствовало расшифровке их первичной структуры и макромолекулярной организации.
Большое значение для развития молекулярной биологии вообще и генетики человека, в особенности, имело открытие Л. Полингом (1940) ненормального гемоглобина S, выделенного из эритроцитов людей с тяжелой наследственной болезнью - серповидно-клеточной анемией. В 1955 - 1957 гг. В. Ингрэм использовал разработанный Ф. Сенгером метод "отпечатков пальцев" (пятен, образуемых отдельными пептидами при хроматографии на бумаге) для анализа продуктов гидролиза гемоглобина S щелочью и трипсином. В 1961 г. Ингрэм сообщил, что гемоглобин S отличается от нормального гемоглобина только по природе одного аминокислотного остатка: в нормальном гемоглобине в седьмом положении цепи находится остаток глютаминовой кислоты, а в гемоглобине S - остаток валина. Тем самым полностью подтвердилось (1949) предположение Полинга, что серповидно-клеточная анемия является болезнью молекулярной природы. Наследуемое изменение всего одного остатка аминокислоты в каждой половинке макромолекулы гемоглобина приводит к тому, что гемоглобин утрачивает способность легко растворяться при низкой концентрации кислорода и начинает кристаллизоваться, что приводит к нарушению структуры клетки. Эти исследования со всей очевидностью показали, что структура белка представляет собой строго определенную аминокислотную последовательность, которая закодирована в геноме. Об исключительном значении первичной структуры белка в формировании уникальной биологически активной конформации макромолекулы свидетельствовали работы К. Анфинсена (1951). Анфинсен показал, что утрачиваемая в результате восстановления биологически активная макроструктура панкреатической рибонуклеазы предопределена аминокислотной последовательностью и может вновь возникать спонтанно при окислении SH-групп остатков цистеина с образованием дисульфидных сшивок в строго определенных местах пептидной цепи фермента.
К настоящему времени детально изучен механизм действия большого числа ферментов и определена структура многих белков.
В 1953 г. Ф. Сенгер установил аминокислотную последовательность инсулина. :Этот белок состоит из двух полипептидных цепей, соединенных двумя дисульфидными сшивками. Одна из цепей содержит всего 21 аминокислотный остаток, а другая - 30 остатков. На расшифровку строения этого сравнительно простого белка Сенгер потратил около 10 лет. В 1958 г. за это выдающееся исследование ему была присуждена Нобелевская премия. После создания В. Стейном и С. Муром (1957) автоматического анализатора аминокислот, идентификация продуктов частичного гидролиза белков значительно ускорилась. В 1960 г. Стейн и Мур уже сообщили о том. что им удалось определить последовательность рибонуклеазы, пептидная цепочка которой представлена 124 аминокислотными остатками. В том же году в лаборатории Г. Шрамма в Тюбингене (ФРГ) Ф. Андерер и другие определили аминокислотную последовательность в белке ВТМ. Затем аминокислотная последовательность была определена в миоглобине (А. Эдмунсон) и α- и β-цепях гемоглобина человека (Г. Браунитцер, Э. Шредер и др.), лизоциме из белка куриного яйца (Ж. Жолле, Д. Кейфилд). В 1963 г. Ф. Шорм и Б. Кейл (ЧССР) установили последовательность аминокислот в молекуле химотрипсиногена. В том же году была определена аминокислотная последовательность трипсиногена (Ф. Шорм, Д. Уолш). В 1965 г. К. Такахаши установил первичную структуру рибонуклеазы T1. Затем последовательность аминокислот была определена еще у нескольких белков.
Как известно, окончательным доказательством правильности определения той или иной структуры является ее синтез. В 1969 г. Р. Мерифилд (США) впервые осуществил химический синтез панкреатической рибонуклеазы. При помощи разработанного им метода синтеза на твердофазовом носителе Мерифилд присоединял к цепочке одну аминокислоту за другой в соответствии с той последовательностью, которая была описана Стейном и Муром. В результате он получил белок, который по своим качествам был идентичен панкреатической рибонуклеазе А. За раскрытие строения рибонуклеазы В. Стейну, С. Муру и К. Анфинсену была в 1972 г. присуждена Нобелевская премия. Этот синтез природного белка открывает грандиозные перспективы, указывая на возможность создания любых белков в соответствии с заранее запланированной последовательностью.
Из рентгеноструктурных исследований У. Астбери (1933) следовало, что пептидные цепи белковых молекул скручены или уложены каким-то строго определенным образом. Начиная с этого времени, многие авторы высказывали различные гипотезы о способах укладки белковых цепей, но до 1951 г. все модели оставались умозрительными построениями, не отвечавшими экспериментальным данным. В 1951 г. Л. Полинг и Р. Кори опубликовали серию блестящих работ, в которых окончательно была сформулирована теория вторичной структуры белков - теория α-спирали. Наряду с этим стало также известно, что белки обладают еще третичной структурой: α-спираль пептидной цепи может быть определенным образом сложена, образуя довольно компактную структуру.
В 1957 г. Дж. Кендрю и его сотрудники впервые предложили трехмерную модель структуры миоглобина. Эта модель затем уточнялась в течение нескольких лет, пока в 1961 г. не появилась итоговая работа с характеристикой пространственной структуры этого белка. В 1959 г. М. Перутц и сотрудники установили трехмерную структуру гемоглобина. На эту работу исследователи затратили более 20 лет (первые рентгенограммы гемоглобина были получены Перутцем в 1937 г.). Поскольку молекула гемоглобина состоит из четырех субъединиц, то, расшифровав его организацию, Перутц тем самым впервые описал четвертичную структуру белка. За работы по определению трехмерной структуры белков Кендрю и Перутцу в 1962 г. была присуждена Нобелевская премия.
Создание Перутцем пространственной модели структуры гемоглобина ПОЗВОЛИЛО. приблизиться к пониманию механизма функционирования этого белка, который, как известно, осуществляет перенос кислорода в животных клетках. Еще в 1937 г. Ф. Гауровиц пришел к выводу о том, что взаимодействие гемоглобина с кислородом, воздуха должно сопровождаться изменением структуры белка. В 60-х годах Перутц и его сотрудники обнаружили заметное смещение цепей гемоглобина после его окисления, вызывавшееся сдвигом атомов железа в результате связывания с кислородом. На этой основе сформировались представления о "дыхании" белковых макромолекул.
В 1960 г. Д. Филлипс и его сотрудники начали рентгеноструктурные исследования молекулы лизоцима. К 1967 г. им более или менее точно удалось установить детали организации этого белка и локализацию отдельных атомов в его молекуле. Кроме этого, Филлипс выяснил характер присоединения лизоцима к субстрату (триацетилглюкозамину). Это позволило воссоздать механизм работы этого фермента. Таким образом, знание первичной структуры и макромолекулярной организации дало возможность не только установить природу активных центров многих ферментов, но и полностью раскрыть механизм функционирования этих макромолекул.
Использование методов электронной микроскопии помогло раскрыть принципы макромолекулярной организации таких сложных белковых образований, как нити коллагена, фибриногена, сократительных фибрилл мышц и др. В конце 50-х годов были предложены модели мышечного сократительного аппарата. Исключительное значение для понимания механизма мышечного сокращения имело открытие В. А. Энгельгардтом и М. Н. Любимовой (1939) АТФ-азной активности миозина. Это означало, что в основе акта мышечного сокращения лежит изменение физико-химических свойств и макромолекулярной организации сократительного белка под влиянием аденозинтрифосфорной кислоты (см. также главу 11).
Для понимания принципов сборки биологических структур существенное значение имели вирусологические исследования (см. главу 25).
Нерешенные проблемы
Основные успехи в современной молекулярной биологии достигнуты в основном в результате изучения нуклеиновых кислот. Тем не менее даже в этой области еще далеко не все проблемы разрешены. Больших усилий потребует, в частности, расшифровка всей нуклеотидной последовательности генома. Эта проблема в свою очередь неразрывно связана с проблемой гетерогенности ДНК и требует разработки новых совершенных методов фракционирования и выделения индивидуальных молекул из суммарного генетического материала клетки.
До сих пор усилия в основном были сосредоточены на раздельном изучении белков и нуклеиновых кислот. В клетке же эти биополимеры неразрывно связаны друг с другом и функционируют главным образом в форме нуклеопротеидов. Поэтому сейчас с особой остротой проявилась необходимость изучения взаимодействия белков и нуклеиновых кислот. На первый план выдвигается проблема узнавания белками определенных участков нуклеиновых кислот. Уже наметились шаги к изучению такого взаимодействия этих биополимеров, без которого немыслимо полное понимание структуры и функций хромосом, рибосом и других структур. Без этого невозможно также уяснить регуляцию активности генов и окончательно расшифровать принципы работы белоксинтезирующих механизмов. После работ Жакоба и Моно появились некоторые новые данные о регуляторном значении мембран в синтезе ядерного материала. Это ставит задачу более глубокого исследования роли мембран в регуляции репликации ДНК. В целом проблема регуляции активности генов и клеточной активности вообще стала одной из важнейших проблем современной молекулярной биологии.
Современное состояние биофизики
В тесной связи с проблемами молекулярной биологии шло развитие биофизики. Интерес к этой области биологии стимулировался, с одной стороны, необходимостью всестороннего изучения действия на организм различного рода излучений, с другой - потребностью исследования физических и физико-химических основ жизненных явлений, протекающих на молекулярном уровне.
Получение точных сведений о молекулярных структурах и совершающихся в них процессах стало возможным в результате применения новых тонких физико-химических методов. На основе достижений электрохимии удалось усовершенствовать метод измерения биоэлектрических потенциалов, применив ионно-избирательные электроды (Г. Эйзенман, Б. П. Никольский, Кхури, 50 - 60-е годы). Все шире входит в практику инфракрасная спектроскопия (с использованием лазерных устройств), позволяющая исследовать конформационные изменения белков (И. Плотников, 1940). Ценные сведения дает также метод электронного парамагнитного резонанса (Е. К. Завойский, 1944) и биохемолюминесцентный метод (Б. Н. Тарусов и др., 1960), которые позволяют, в частности, судить о транспорте электронов при окислительных процессах.
К 50-м годам биофизика завоевывает уже прочное положение. Возникает потребность в подготовке квалифицированных специалистов. Если в 1911 г. в Европе только в университете г. Печ, в Венгрии, была кафедра биофизики, то к 1973 г. такие кафедры существуют почти во всех крупных университетах.
В 1960 г. было организовано Международное общество биофизиков. В августе 1961 г. состоялся первый Международный биофизический конгресс в Стокгольме. Второй конгресс был проведен в 1965 г. в Париже, третий - в 1969 г. в Бостоне, четвертый - в 1972 г. в Москве.
В биофизике сохраняется четкое разграничение между двумя различными по содержанию направлениями - молекулярной биофизикой и клеточной биофизикой. Это разграничение получает и организационное выражение: создаются раздельные кафедры этих двух направлений биофизики. В Московском университете первая кафедра биофизики была создана в 1953 г. на биолого-почвенном факультете, несколько позже возникла кафедра биофизики на физическом факультете. По такому же принципу организовывались кафедры во многих других университетах.
Молекулярная биофизика
В последние годы все больше укреплялась связь молекулярной биофизики с молекулярной биологией, и сейчас бывает иногда трудно определить, где проходит граница раздела между ними. В генеральном наступлении на проблему наследственной информации такая кооперация биофизики с молекулярной биологией неизбежна.
Главным направлением в исследовательской работе является изучение физики нуклеиновых кислот - ДНК и РНК. Применение указанных выше методов и прежде всего рентгеноструктурного анализа способствовало расшифровке молекулярной структуры нуклеиновых кислот. В настоящее время ведутся интенсивные исследования по изучению поведения этих кислот в растворах. Особое внимание уделяется при этом конформационным переходам "спираль-клубок", изучаемым по изменениям вязкости, оптическим и электрическим показателям. В связи с изучением механизмов мутагенеза развиваются исследования по изучению действия ионизирующей радиации на поведение нуклеиновых кислот в растворах, а также действия радиации на нуклеиновые кислоты вирусов и фагов. Всестороннему анализу подвергалось влияние ультрафиолетового излучения, некоторые спектральные участки которого, как известно, хорошо поглощаются нуклеиновыми кислотами. Большой удельный вес в такого рода исследованиях занимает обнаружение активных радикалов нуклеиновых кислот и белков методом электронного парамагнитного резонанса. С применением этого метода связано возникновение целого самостоятельного направления.
Проблема кодирования информации ДНК и РНК и ее передачи при синтезе белка давно интересовала молекулярную биофизику, и физики неоднократно высказывали по этому поводу те или иные соображения (Э. Шредингер, Г. Гамов). Расшифровка генетического кода вызвала многочисленные теоретические и экспериментальные исследования по структуре спирали ДНК, механизму скольжения и закручивания ее нитей, по изучению физических сил, участвующих в данных процессах.
Значительную помощь молекулярная биофизика оказывает молекулярной биологии в изучении структуры белковых молекул при помощи рентгеноструктурного анализа, впервые примененного в 1930 г. Дж. Берналом. Именно в результате использования физических методов в сочетании с биохимическими (ферментативные методы) была вскрыта молекулярная конформация и последовательность расположения аминокислот в ряде белков.
Современные электронно-микроскопические исследования, выявившие наличие в клетках и ее органоидах сложных мембранных систем, стимулировали попытки понять их молекулярное строение (см. главы 10 и 11). Изучается прижизненно химический состав мембран и, в частности, свойства их липидов. Было выяснено, что последние способны к переокислению и неферментативным реакциям цепного окисления (Ю. А. Владимиров и Ф. Ф. Литвин, 1959; Б. Н. Тарусов и др., 1960; И. И. Иванов, 1967), приводящим к нарушению мембранных функций. Для изучения состава мембран стали пользоваться также методами математического моделирования (В. Ц. Пресман, 1964 - 1968; М. М. Шемякин, 1967; Ю. А. Овчинников, 1972).
Клеточная биофизика
Знаменательным событием в истории биофизики явилось формирование в 50-х годах четких представлений о термодинамике биологических процессов, в результате чего окончательно отпали предположения о возможности самостоятельного образования энергии в живых клетках вопреки второму закону термодинамики. Понимание действия этого закона в биологических системах связано с введением бельгийским ученым И. Пригожиным (1945) * в биологическую термодинамику понятия открытых систем, обменивающихся с внешней средой энергией и материей. Пригожин показал, что положительная энтропия образуется в живых клетках при рабочих процессах соответственно второму закону термодинамики. Введенные им уравнения определили условия, при которых возникает так называемое стационарное состояние (ранее его именовали также динамическим равновесием), при котором количество свободной энергии (негэнтропии), поступающей в клетки с пищей, компенсирует ее расход, а положительная энтропия выводится. Это открытие подкрепило общебиологическую идею о неразрывной связи внешней и внутренней среды клеток. Оно положило начало реальному изучению термодинамики живы" систем, в том числе методом моделирования (А. Бэртон, 1939; А. Г. Пасынский, 1967).
* (Общую теорию открытых систем впервые выдвинул Л. Берталанфи в 1932 г. )
Согласно основному принципу биотермодинамики, необходимым условием существования жизни оказывается стационарность в развитии ее биохимических процессов, для осуществления которой необходима координация скоростей многочисленных реакций обмена веществ. На основе новой биофизической термодинамики возникло направление, выделяющее внешние и внутренние факторы, которые обеспечивают эту координацию реакций и делают ее устойчивой. За последние два десятилетия выявлена большая роль в поддержании стационарного состояния системы ингибиторов и особенно антиоксидантов (Б. Н. Тарусов и А. И. Журавлев, 1954, 1958). Установлено, что надежность стационарного развития связана с факторами внешней среды (температурой) и физико-химическими свойствами среды клеток.
Современные принципы биотермодинамики позволили дать физико-химическое истолкование механизму адаптации. По нашим данным, приспособление к условиям внешней среды может происходить только в том случае, если при их изменении организм способен установить стационарность в развитии биохимических реакций (Б. Н. Тарусов, 1974). Встал вопрос о разработке новых методов, которые позволили бы оценивать стационарное состояние прижизненно и прогнозировать его возможные нарушения. Большую пользу сулит внедрение в биотермодинамику и исследования процессов биологической адаптации кибернетических принципов саморегулирующихся систем. Стало ясно, что для решения вопроса об устойчивости стационарного состояния важен учет так называемых возмущающих факторов, к которым относятся, в частности, неферментативные реакции окисления липидов. В последнее время все более расширяются исследования процессов переокисления в липидных фазах живых клеток и нарастания активных радикальных продуктов, нарушающих регуляторные функции мембран. Источником информации об этих процессах служит как обнаружение активных перекисных радикалов, так и перекисных соединений биолипидов (А. Таппель, 1965; И. И. Иванов, 1965; Е. Б. Бурлакова, 1967 и другие). Для обнаружения радикалов используют биохемолюминесценцию, возникающую в липидах живых клеток при их рекомбинации.
На основе физико-химических представлений о стабильности стационарного состояния возникли биофизические представления об адаптации растений к изменениям условий внешней среды как нарушении ингибирующих антиокислительных систем (Б. Н. Тарусов, Я. Е. Доскоч, Б. М. Китлаев, А. М. Агавердиев, 1968 - 1972). Это открыло возможность оценивать такие свойства, как морозоустойчивость и солеустойчивость, а также делать соответствующие прогнозы при селекции сельскохозяйственных растений.
В 50-х годах было открыто сверхслабое свечение - биохемолюминесценция ряда биологических объектов в видимой и инфракрасной частях спектра (Б. Н. Тарусов, А. И. Журавлев, А. И. Поливода). Это стало возможным в результате разработки методов регистрации сверхслабых световых потоков при помощи фотоэлектронных умножителей (Л. А. Кубецкий, 1934). Являясь результатом биохимических реакций, протекающих в живой клетке, биохемолюминесценция позволяет судить о важных окислительных процессах в цепях переноса электронов между ферментами. Открытие и изучение биохемолюминесценции имеет большое теоретическое и практическое значение. Так, Б. Н. Тарусов и Ю. Б. Кудряшов отмечают большую роль продуктов окисления ненасыщенных жирных кислот в механизме возникновения патологических состояний, развивающихся под действием ионизирующих излучений, при канцерогенезе и других нарушениях нормальных функций клетки.
В 50-х годах в связи с бурным развитием ядерной физики из биофизики выделилась радиобиология, исследующая биологическое действие ионизирующих излучений. Получение искусственных радиоактивных изотопов, создание термоядерного оружия, атомных реакторов и развитие других форм практического использования атомной энергии поставило со всей остротой проблему защиты организмов от вредного действия ионизирующей радиации, разработки теоретических основ профилактики и лечения лучевой болезни. Для этого необходимо было в первую очередь выяснить, какие компоненты клетки и звенья обмена веществ наиболее уязвимы.
Объектом изучения биофизики и радиобиологии стало выяснение природы первичных химических реакций, возникающих в живых субстратах под воздействием энергии излучений. Здесь было важно не только понять механизмы этого явления, но и суметь воздействовать на процесс размена физической энергии на химическую, уменьшить его коэффициент "полезного" действия. Работам в этом направлении положили начало исследования школы Н. Н. Семенова (1933) в СССР и Д. Хиншельвуда (1935) в Англии.
Большое место в радиобиологических исследованиях заняло изучение степени радиационной сопротивляемости различных организмов. Было установлено, что повышенная радиорезистентность (например, грызунов пустынь) обусловлена высокой антиокислительной активностью липидов клеточных мембран (М. Чанг и др., 1964; Н. К. Огрызов и др., 1969). Оказалось, что в формировании антиоксидативных свойств этих систем большую роль играют токоферолы, витамин К и тиосоединения (И. И. Иванов и др., 1972). В последние годы большое внимание привлекают к себе также исследования механизмов мутагенеза. С этой целью изучается действие ионизирующих излучений на поведение нуклеиновых кислот и белков in vitro, а также в вирусах и фагах (А. Густафсон, 1945 - 1950).
Борьба за дальнейшее повышение эффективности химической защиты, поиск более эффективных ингибиторов и принципов ингибирования остаются в этом направлении основными задачами биофизики.
Продвинулось вперед исследование возбужденных состояний биополимеров, определяющих их высокую химическую активность. Наиболее успешно шло изучение возбужденных состояний, возникающих на первичной стадии фотобиологических процессов - фотосинтеза и зрения.
Так, сделан солидный вклад в понимание первичной активации молекул пигментных систем растений. Установлено большое значение переброски (миграции) энергии возбужденных состояний без потерь с активированных пигментов на другие субстраты. Большую роль в развитии этих представлений сыграли теоретические работы А. Н. Теренина (1947 и позднее). А. А. Красновский (1949) открыл и исследовал реакцию обратимого фотохимического восстановления хлорофилла и его аналогов. Ныне складывается всеобщее убеждение, что в ближайшем будущем можно будет воспроизвести фотосинтез в искусственных условиях (см. также главу 5).
Биофизики продолжают работать над раскрытием природы мышечного сокращения и механизмов нервного возбуждения и проведения (см. главу 11). Актуальное значение приобрели также исследования механизмов перехода от возбужденного состояния к норме. Возбужденное состояние рассматривают теперь как результат автокаталитической реакции, а торможение - как следствие резкой мобилизации ингибиторной антиокислительной активности в результате молекулярных перегруппировок в таких соединениях, как токоферол (И. И. Иванов, О. Р. Кольс, 1966; О. Р. Кольс, 1970).
Важнейшей общей проблемой биофизики остается познание качественных физико-химических особенностей живой материи. Такие свойства, как способность живых биополимеров избирательно связывать калий или поляризовать электрический ток, не удается сохранить даже при их самом осторожном извлечении из организма. Поэтому клеточная биофизика продолжает интенсивно разрабатывать критерии и методы для прижизненного исследования живой материи.
Несмотря на молодость молекулярной биологии, успехи, достигнутые ею в этой области, поистине ошеломляющи. За сравнительно короткий срок установлена природа гена и основные принципы его организации, воспроизведения и функционирования. Более того, осуществлено не только размножение генов in vitro, но и впервые завершен полный синтез самого гена. Полностью расшифрован генетический код и разрешена важнейшая биологическая проблема специфичности биосинтеза белка. Выявлены и исследованы главные пути и механизмы образования белка в клетке. Полностью определена первичная структура многих транспортных РНК - специфических молекул-адапторов, осуществляющих перевод языка нуклеиновых матриц на язык аминокислотной последовательности синтезирующегося белка. До конца расшифрована аминокислотная последовательность многих белков и установлена пространственная структура некоторых из них. Это дало возможность выяснять принцип и детали функционирования молекул ферментов. Осуществлен химический синтез одного из ферментов - рибонуклеазы. Установлены основные принципы организации разных субклеточных частиц, многих вирусов и фагов и разгаданы основные пути их биогенеза в клетке. Вскрыты подходы к пониманию путей регуляции активности генов и выяснению регуляторных механизмов жизнедеятельности. Уже простой перечень этих открытий свидетельствует о том, что вторая половина XX в. ознаменовалась огромным прогрессом биологии, который обязан прежде всего углубленному изучению структуры и функций биологически важнейших макромолекул - нуклеиновых кислот и белков.
Достижения молекулярной биологии уже сегодня используются на практике и приносят ощутимые плоды в медицине, сельском хозяйстве и некоторых отраслях промышленности. Несомненно, что отдача этой науки будет возрастать с каждым днем. Однако главным итогом все же следует считать, что под влиянием успехов молекулярной биологии укрепилась уверенность в существовании неограниченных возможностей на пути раскрытия самых сокровенных тайн жизни.
В будущем, по-видимому, будут открыты новые пути исследования биологической формы движения материи - с молекулярного уровня биология перейдет на атомарный уровень. Однако сейчас не найдется, пожалуй, ни одного исследователя, который мог бы достаточно реально предсказать развитие молекулярной биологии даже на ближайшие 20 лет.
31.2
Для друзей!
Справка
Молекулярная биология выросла из биохимии в апреле 1953 года. Ее появление связано с именами Джеймса Уотсона и Френсиса Крика, которые открыли структуру молекулы ДНК. Открытие стало возможным благодаря исследованию генетики, бактерий и биохимии вирусов. Профессия молекулярный биолог не имеет широкого распространения, но на сегодняшний день ее роль в современном обществе очень велика. Большое количество заболеваний, в том числе, проявляющихся на генетическом уровне, требует от ученых поиска решений данной проблемы.
Описание деятельности
Вирусы и бактерии постоянно мутируют, а это значит, что человеку перестают помогать лекарства и болезни становятся трудноизлечимыми. Задача молекулярной биологии - опередить этот процесс и разработать новое средство от болезней. Ученые работают по отлаженной схеме: блокировка причины заболевания, устранение механизмов наследственности и облегчение тем самым состояния пациента. В мире существует ряд центров, клиник и больниц, где молекулярные биологи в помощь пациентам разрабатывают новые способы лечения.
Трудовые обязанности
В обязанности молекулярного биолога входит изучение процессов внутри клетки (например, изменения в ДНК при развитии опухолей). Также специалисты изучают особенности ДНК, их влияние на целый организм и отдельную клетку. Такие исследования проводятся, например, на основании ПЦР (полимеразная цепная реакция), которая позволяет сделать анализ организма на инфекции, наследственные заболевания и определить биологическое родство.
Особенности карьерного роста
Профессия молекулярный биолог довольно перспективна в своей сфере и уже сегодня претендует на первые места в рейтинге медицинских профессий будущего. Кстати, молекулярному биологу не обязательно все время оставаться в этой сфере. Если возникнет желание сменить род занятий, он может переквалифицироваться в менеджеры по продажам лабораторного оборудования, начать разрабатывать приборы для различных исследований или открыть свое дело.